腺苷二磷酸葡萄糖焦磷酸化酶(ADP-glucose pyrophosphorylase,AGP)活性测定试剂盒说明书
生物化学-名词英汉互译

英汉互译1.RDA:核糖核酸2.DNA:脱氧核糖核酸3.“m”(methy-):甲基化修饰基团4.bp:碱基对5.NMP:核苷一磷酸NDP:核苷二磷酸NTP:核苷三磷酸(N表示A、T、C、G)6.AMP:腺苷酸GMP:鸟苷酸CMP:胞苷酸UMP:尿苷酸7.dAMP:脱氧腺苷酸dGMP:脱氧鸟苷酸8.ADP:腺苷二磷酸ATP:腺苷三磷酸9.cAMP:3′、5′—环腺苷酸 cGMP:3′、5′—环鸟苷酸10.snRNA:核内小RNA snoRNA:核仁小RNA miRNA:微小RNA11.H-DNA:三链DNA C结构:共价闭合环状结构13.mRNA:信使RNA tRNA:转运RNA rRNA:核糖体RNA14.DHU:二氢尿嘧啶 15.Tm:熔点或熔解温度16.hnRNA:核内不均一RNA polyA:多聚核苷酸17.Ala(A):丙氨酸 18.Gly(G):甘氨酸 19.Asp(D) :天冬氨酸Val(V):缬氨酸Ser(S):丝氨酸Glu(E):谷氨酸Lev(L):亮氨酸Thr(T):苏氨酸Lys(K):赖氨酸Ile(I):异亮氨酸Cys(C):半胱氨酸Arg(R):精氨酸Pro(P):脯氨酸Tyr(Y):络氨酸His(H):组氨酸Phe(F):苯丙氨酸Asn(N):天冬酰胺Trp(W):色氨酸Gln(Q):谷氨酰胺Met(M):甲硫氨酸20.TRH:促甲状腺素释放因子22.LRF:促黄体生成激素释放因子23.GRIF:生长激素释放抑制因子24.ADH:加压素25.ACTH:促肾上腺皮质激素26.βMSH:促黑激素β27.ANP:心钠肽28.NPY:神经肽29.IUPA:应用化学联合会1-纤维素:羧甲基纤维素31.DEAF纤维素:二乙氨基纤维素32.SDS:十二烷基酸钠33.EC:国际酶学委员会34.DIPF:二异丙基氟磷酸35.NAG:N-乙酰葡胺36.NAM:N-乙酰胞壁酸37.Km:米式常数38.EDTA:乙二胺四乙酸39.AT Case:天冬氨酸转氨甲酰酶40.LDH:乳酸脱氢酶41.ADH:乙酸脱氢酶42.Kcat:酶的催化常数(酶的转换数)43.PAGE:聚丙酰胺凝胶电泳44.TPP:焦磷酸硫胺素45.FMN:黄素单核苷酸46.FAD:黄素腺嘌呤二核苷酸47.NAD:烟酰胺腺嘌呤二核苷酸48.NADP:烟酰胺腺嘌呤二核苷酸磷酸49.DHFA:二氢叶酸50.THFA:四氢叶酸51.GSH:谷胱甘肽52.FA:脂肪酸53.PG:前列腺素54.SQDG:6-硫酸D-异鼠李糖二酰甘油55.PC:磷脂酰胆碱PE:磷脂酰胆胺PS:磷脂酰丝氨酸PI:磷脂酰肌醇56.MGDG:单半乳糖二酰甘油DGDG:双乳糖二酰甘油57.PA:磷脂酸PG:磷脂酰甘油58.CL(DPG):心磷脂59.EMP途径:糖酵解途径60.FBP:1,6-二磷酸果糖61.DPGA:1,3-二磷酸甘油酸62.PEP:磷酸烯醇式丙酮酸63.PFK:磷酸果糖激酶64.TCA循环:三羧酸循环65.PPP途径:磷酸戊糖途径66.ETS:电子传递链(呼吸链)67.UQ(CoQ):泛醌(辅酶Q)68.OSCP:寡酶素敏感性蛋白69.SOD:超氧物歧化酶70.CAT:过氧化氢酶71.PC:电子载体质体蓝素Fd:铁氧化还原蛋白Fp:还原酶71.P SⅠ:光系统Ⅰ PSⅡ:光系统Ⅱ72.A:原初电子受体D:原初电子供体73.cyt:细胞色素74.UDPG:尿苷二磷酸葡萄糖ADPG:腺苷二磷酸葡萄糖75.TP:磷酸丙糖76.FBPase:果糖-1,6-二磷酸酶77.PFK:磷酸果糖激酶78.PFP:磷酸果糖焦磷酸化酶79.GAC:乙醛酸循环80.BCCP:生物素羧基载体蛋白81.BC:生物素羧化酶82.CT:羧基转移酶83.ACP:脂酰基载体蛋白84.FAS:脂肪酸合酶85.MV A:3-甲基-3,5-二羟戊酸(甲瓦龙酸)86.β-羟-β-甲基戊二酸单酰CoA:(HMG-CoA)87.IPP:异戊烯醇焦磷酸酯DPP:3,3-二甲基丙烯焦磷酸酯GPP:()牛儿焦磷酸酯FPP:法呢焦磷酸酯88.SCP:固体载体蛋白89.DIPF:二丙基氟磷酸90.GPT:谷丙转氨酶GOT:谷草转氨酶91.Fd:还原型铁氧还蛋白92.OAA:草酰乙酸93.APS:腺苷酰硫酸94.PAPS:磷酸腺苷酰硫酸95.DNase:脱氧核糖核酸酶RNase:核糖核酸酶96.PRPP:5-磷酸核糖焦磷酸97.XMP:黄嘌呤核苷酸98.OPM:乳清酸核苷酸99.CP SⅡ:氨甲酰磷酸合成酶Ⅱ100.HGPRT:次黄嘌呤-鸟嘌呤磷酸核糖转移酶101.PCR:聚合酶链反应102.PDI:蛋白质二硫键异构酶103.SRP:信号肽识别颗粒104.:RTPK:受体型络氨酸蛋白激酶105.SR:固醇受体AR:雄性激素受体GR:糖皮质激素受体ER:雌性激素受体106.NAD+:氧化态的辅酶Ⅰ107.NADH:还原态辅酶ⅠNADP+:氧化肽的辅酶ⅡNADPH:还原态辅酶Ⅱ。
初中化学 酶 腺苷二磷酸 知识点总结

初中化学酶腺苷二磷酸知识点总结
一、酶的基本概念和作用:
- 酶是一种生物催化剂,能够加速化学反应的进行。
- 酶能够在生物体内促进新陈代谢的进行,维持生命活动。
- 酶能够降低反应的活化能,使反应更容易进行。
二、酶的特点:
- 酶是一种蛋白质,由一条或多条多肽链构成。
- 酶具有高度的专一性,只催化特定的底物反应。
- 酶在反应中会发生构象变化,但不参与反应本身。
三、腺苷二磷酸(ADP)的作用:
- 腺苷二磷酸是一种细胞内能量转化的物质。
- 当供能反应进行时,三磷酸腺苷(ATP)会转变为腺苷二磷酸(ADP)和无机磷酸。
- ADP能够通过再次与无机磷酸结合,重新转变为ATP,供给生物体所需的能量。
四、ADP与能量转化的关系:
- ADP和ATP之间的转化过程是细胞内能量转化的关键环节。
- 当细胞需要能量时,ADP会通过与无机磷酸结合形成ATP,
并释放出能量。
- 当细胞释放能量时,ATP会通过水解反应转变为ADP和无机磷酸,并释放出能量。
以上是初中化学中关于酶和腺苷二磷酸的一些基本知识点总结。
对于更详细的内容,建议参考相关教材或资料。
二氧化碳人工合成淀粉流程

二氧化碳人工合成淀粉流程生物界对于糖类的摄取与利用是其运转的重要组成部分,淀粉作为该过程的代表性物质,吸引了科学界的广泛关注。
生物摄取糖类,将其不断分解成各种小分子,这些被分解的物质中,二氧化碳和水居多。
近日,我国中科院科研团队通过“光能-电能-化学能”的能量转变,成功构建了从二氧化碳人工合成淀粉仅需十一步反应的途径!1.第一步:二氧化碳与氢气在氧化锌-二氧化锆的催化作用下反应生成甲醇。
其中氧化锌-二氧化锆催化剂也是该科研团队在此领域的一大重要研发产物。
2.第二步:甲醇在过氧化氢作用下氧化为甲醛。
前两步的反应非常简单,中学化学知识就已经足够我们理解,而第三步开始就是整个反应的难点与重头戏。
3.第三步;两个甲醛分子在甲醛酶的作用下发生羟醛缩合,生成二羟基丙酮。
在这个过程中,一个甲醛分子A断开一个C-H键,另一个甲醛分子B断开一C=O 键变成C-O键,然后A的碳原子与B的碳原子间形成C-C键,A上游离出的氢原子与B的氧原子间形成O-H键。
同理,在A的另一侧也会发生该反应。
这一步的反应,将一碳化合物转化为了三碳化合物。
4.第四步:二羟基丙酮在二羟基丙酮激酶的催化下与ATP(三磷酸腺苷)发生反应生成磷酸二羟基丙酮。
5.第五步:将磷酸二羟基丙酮转化为它的同分异构体磷酸甘油醛。
这个过程中,磷酸丙糖异构酶将从C端碳上拆下来一个氢原子,最后磷酸丙糖异构酶又会将这个氢原子还给中间的碳原子。
6.第六步:将第五步得到的磷酸甘油醛与第四步得到的磷酸二羟基丙酮进行成环反应得到1,6-二磷酸果糖。
这一步的反应较复杂,继三碳化合物阶段后进入了六碳化合物阶段。
7.第七步:将1,6-二磷酸果糖的1号碳位上的磷酸根换为羟基。
8.第八步:磷酸葡萄糖异构酶将6-磷酸果糖变成了6-磷酸葡萄糖。
目前,第七步反应产物的五元环变成了六元环。
9第九步:在磷酸葡萄糖变位酶的作用下磷酸根从1号碳上转移至6号碳上,与6号碳中羟基的氢原子互换位置。
10.第十步:让第九步的产物在腺苷二磷酸葡萄糖焦磷酸化酶催化下与ATP 反应,使得第九步产物中磷酸根的位置被替换为一个ADP。
小麦蔗糖合成酶和腺苷二磷酸葡萄糖焦磷酸化酶研究进展

小麦蔗糖合成酶和腺苷二磷酸葡萄糖焦磷酸化酶的研究进展作物杂志Crops2008.6小麦蔗糖合成酶和腺苷二磷酸葡萄糖焦磷酸化酶的研究进展术姜丽娜李冬芬李春喜邵云摘要植物蔗糖合成酶(SS)和腺苷二磷酸葡萄糖焦磷酸化酶(AGPase)在植物淀粉生物合成过程中起着重要的作用,淀粉是小麦等禾谷类作物子粒胚乳的重要组成成分,与作物的产量和品质密切相关.综述了小麦蔗糖合成酶和腺苷二磷酸葡萄糖焦磷酸化酶的细胞定位生物学功能,分子生物学分析及酶活性的调控,并进一步对两种酶的研究作出设想.关键词小麦;蔗糖合成酶;腺苷二磷酸葡萄糖焦磷酸化酶蔗糖合成酶(ss)是蔗糖代谢关键酶中极其重要的一种酶,催化反应:尿苷二磷酸葡萄糖(UDPG)+作者简介:姜丽娜,副教授,主要从事小麦栽培生理研究工作,河南师范大学生命科学学院,453007,河南新乡李冬芬,李春喜,邵云,通讯地址同第1作者基金项目:”十一五”国家科技支撑计划(2006BAI)o2AI5,2OO6BAK02A25)资助收稿日期:2008—08—05:修回日期:2008—10—28果糖一蔗糖+尿苷二磷酸(UDP).蔗糖合成最适pH值8.0~9.5,蔗糖裂解最适pH值5.5~6.5.在小麦旗叶中,ss参与蔗糖的合成,在子粒中,ss的作用主要是催化子粒中蔗糖的降解,为淀粉合成提供前提物质.AGPase催化1一磷酸葡萄糖(G1P)与三磷酸腺苷(ATP)反应形成ADPG,其作为合成淀粉的直接底物,是子粒发育过程中控制淀粉含量的酶.AGPase表达受抑制或过量表达会引起淀粉含量的下降或增加,本文就近年来有关两种酶的研究进展做一介绍.l酶的功能及细胞定位1.1蔗糖合成酶蔗糖合成酶是由Cardini等于1955年首次在小麦胚芽中发现.目前国外研究者已成功地将sS基因从小麦中克隆出来j,随着研究的不断深入,已知ss在小麦等作物产量调控方面具有重要的ResearchAdvancesonthePigmentDegradationand ItsRegulationMeasuresduringtheFlue--curedTobaccoCuringSongZhaopeng,JingYongfeng,LiChangjun3,QiYongjie4,ZhangQinsong3,GongChangrong’(CollegeofAgronomy,HenanAgriculturalUniversity,Zhengzhou450002,Henan;Chinese TobaccoIndustrialCorporationofHunan,Changsha410000,Hunan;FengjieTobaccoCompanyofChongqing,Fengjie40460 0,Chongqing;ChineseTobaccoIndustrialCorporationofGuangxi,Nanning530001,Guangxi,China) AbstractThedegradationofpigmentduringtheflue—curedtobaccocuringhadgreateffects ontobaccoquality.In ordertoexplorepigmentdegradationmechanismduringcuring,understandrelationshipsbet weenthepigmentdegra- tionanditschangesonthetobaccointercellandlookfornewwaystoreducethepigmentconten t,theresearchad—vancesonthetobaccopigmentduringcuringweresummarized,includingthecontentsoftoba ccopigment,thedegra—dationmechanismofitsmajorconstituent,thechangesofpigmentcontentsduringcuringandi tsregulationmeasure.KeywordsFlue-curedtobacco;Curing;Pigment;Regulation112008.6作物杂志Crops作用.SS是一种胞质酶,是促使蔗糖进入各种代谢途径的关键酶之一3j,在大多数植物的贮藏器官中得到明显的表达,其活性通常与淀粉的积累或者器官的生长发育相联系.目前SS已经被确认有两种不同的存在形式SSI和SSI!,大部分以可溶性状态存在于细胞质中,有些不溶性的SS则附着在细胞膜上.SS可催化蔗糖合成与分解,但通常认为sS主要起分解蔗糖的作用,研究表明,其分解产物尿苷二磷酸葡萄糖(UDPG)可为细胞壁构建和糖酵解代谢提供底物,同时也是支链淀粉和直链淀粉合成的前体,淀粉是小麦子粒的主要组成成分,子粒的灌浆充实过程,主要是胚乳中淀粉的合成与积累过程,SS是蔗糖向淀粉转化过程中的第1个限速酶,其活性高低和持续期长短决定着子粒的灌浆速率和持续时间,并与子粒的体积和重量密切相关.李春燕等研究表明,SS活性与蔗糖含量变化及淀粉合成速率变化相吻合,当ss活性处于较高水平时,蔗糖消耗很快,其含量显着降低,而此时也正是淀粉合成的高峰期.王旭东等也曾提到ss是子粒中蔗糖降解的关键酶,与淀粉合成的快慢和粒重呈正相关,SS活性增强,有利于淀粉合成和粒重的提高.因此,子粒中sS调节着淀粉的合成,其活性的高低反映了子粒降解利用蔗糖的能力¨,ss活性高,则合成淀粉的底物就充足.现已证明该酶是小麦子粒淀粉合成的一个调控因子.Wang等0把库器官中SS的活性作为库强度的标志,而谢祝捷等”的分析结果却表明,小麦子粒淀粉产量和含量与蔗糖磷酸合成酶(SPS),可溶性淀粉合成酶(SSS)和淀粉合成酶(GBSS)活性的关系比与SS活性的关系更为密切.因此关于ss在小麦淀粉合成中的地位和影响程度还有待进一步研究.1.2腺苷二磷酸葡萄糖焦磷酸化酶AGPase多存在于植物叶片和贮藏器官中,其在光合细胞中定位于质体内,而在非光合器官中的定位却尚无定论.小麦等禾谷类作物胚乳中有两种不同形式的AGPase,它们分别是胞质的和质体的,其中胞质形式占65%~95%的活性. AGPase在植物体内催化由蔗糖得到的G1P与ATP反应形成腺苷二磷酸葡萄糖(ADPG)并释放Pi,而ADPG作为淀粉合成酶的底物参与直链12淀粉和支链淀粉的合成,是水稻,小麦,玉米,马铃薯等作物淀粉合成的关键酶,其活性大小与淀粉积累速率和灌浆速率呈正相关¨.小麦淀粉合成关键酶活性及其与子粒淀粉积累速率关系的研究也表明,AGPase活性与直链淀粉,支链淀粉,总淀粉的积累速率及灌浆速率呈极显着正相关.于振文等研究表明,开花后7d,14d,21d,28d和35d小麦子粒AGPase活性与各期总淀粉积累速率均呈显着或极显着正相关.1.3SS,AGPase与淀粉合成子粒中淀粉合成的第一步反应是蔗糖在ss的催化下,分解成尿苷二磷酸葡萄糖(UDPG)和果糖(F),然后再经一系列反应生成淀粉,参与淀粉合成的酶很多,如SS,AGPase,淀粉合成酶,淀粉分支酶等,Nakamura等17]认为AGPase和淀粉分支酶是控制淀粉合成的关键酶;Jener等¨认为ss, AGPase均为调节淀粉合成的因子;而梁建生等1,姜东等11¨认为SS,AGPase等是淀粉合成的关键酶;也有研究表明在田间条件下,麦类作物胚乳淀粉合成速率,可能受可溶性淀粉合成酶控制要比受AGPase活性的丧失使子粒灌浆停止有更明确的关系;直链淀粉和支链淀粉合成速率与淀粉合成有关酶活性的相关分析表明,支链淀粉合成速率与SS,AGPase的活性呈极显着正相关, 直链淀粉合成速率与SS,AGPase的活性分别呈极显着正相关,显着相关_】.总之,前人的报道均证明子粒淀粉合成过程主要受SS,AGPase等酶的催化,在子粒淀粉积累过程中,Ss和AGPase均起重要作用,但是对于二者在淀粉合成中作用大小的认识也存在着较大的差异:Preiss等认为AGPase为淀粉生物合成的限速酶,该酶活性的大小直接关系到淀粉合成的速率和最终淀粉合成量的多寡;Okita等则指出,ss活性大小与淀粉积累的量相一致;亦有研究表明在淀粉的生物合成中,AGPase的活性变化对淀粉产量的影响较大,sS 和UDPG焦磷酸化酶也能催化淀粉的合成,李春燕等_8]研究表明,子粒灌浆前期ss起着重要作用,灌浆中期AGPase,SSS起主要作用,灌浆中后期GBSS起主要作用,且子粒AGPase,SSS,GBSS 与子粒灌浆速率,淀粉积累速率达极显着正相关. 但在子粒整个灌浆过程淀粉合成途径中各种酶峰值时间不一致,由此说明SS,AGPase等酶在子粒作物杂志Crops2008.6灌浆过程的不同时期共同协调淀粉的合成进程, 或者说它们对淀粉合成的作用在时间(时期)上可能会有差异.2分子生物学研究2.1蔗糖合成酶SS是相对分子质量为83—100kD的亚基构成的四聚体.一般认为在大多数植物中ss至少有两种同工酶,它们通常都有较高的氨基酸序列同源性和相似性的生化性质,研究了黑麦草,绿竹,菜豆,马铃薯等植物水溶性ss的结构,了解了其核苷酸序列及其推导的氨基酸序列的结构特征,整条肽链的疏水/亲水性,可能的亚细胞定位, 跨膜结构域,蛋白质的二级结构及功能域.但是,目前对小麦SS这方面的研究尚属空白,因此, 用生物信息学方法探明小麦等禾谷类作物SS的高级结构及其决定的潜在功能对了解整个糖代谢意义重大.ss基因的大小约5.9kb,eDNA的长度约2.7kb,编码约820个氨基酸,在单子叶植物中由两个非等位基因Susl和Sus2编码.六倍体小麦的SuslmRNA在缺氧和冷休克(6~C)下迅速增加, 而Sus2mRNA不受影响;对其黄化叶进行光照, SuslmRNA水平明显降低而Sus2mRNA水平增加;SS基因在表达调控上明显不同,基因表达具有发育和组织器官特异性一.目前研究者已深入研究了水稻,拟南芥],玉米],柑橘的基因家族成员,基因的表达,基因的功能.因此,对小麦SS基因家族成员及其表达,SS同工酶的功能的进一步研究将成为今后研究的一个方向.2.2腺苷二磷酸葡萄糖焦磷酸化酶AGPase由两个大亚基(1AGPase)和两个小亚基(SAGPase)组成的异型四聚体,分子量在200~400kD,小麦包含与马铃薯块茎AGPase大小亚基相似大小的肽链,即分子量为51kD的大亚基和50kD的小亚基29j.小麦子粒中AGPase大小亚基相对保守,氨基酸序列同源性大于85%,而大亚基同源性相对较低,但高于物种自身大小亚基序列间的同源性E30].AGPase的每个亚基都由不同的基因编码日¨,且每种亚基都含有酶催化所必需的基团,因而各自都能在缺少另一个亚基的情况下维持酶活性,而两种亚基的共同存在则可协调地进行催化工作.已有学者通过对马铃薯大小亚基eDNA克隆分别或一起在E.coli中表达,从而确定了每个亚基具有的特异性功能L33j.也有多位研究者通过对不同植物大小亚基的重要氨基酸和结构域的点突变研究发现,AGPase小亚基主要起合成作用,大亚基主要起调节作用343.目前,六倍体小麦AGPase的两个亚基已从其发育胚乳的eDNA 文库中获得:Anisworth等一从中国春胚乳的eDNA 文库中筛选到编码该酶小亚基的eDNA克隆,开放读码框为1422bp,推测编码产物的分子量为521kD;用中国春缺体一四体系将该酶小亚基的基因定位在7A,7B和7D染色体上,以单拷贝形式存在,编码由473个氨基酸残基组成的蛋白质,存在一个由22个氨基酸组成的叶绿体/造粉体前导肽;Oliver等E353获得大亚基eDNA克隆,但没有对基因进行定位,由于AGPase在粗提时具有热稳定性,然而要得到高纯度且稳定的酶却十分困难,因此我们可以用上述方法将小麦AGPase大小亚基eDNA克隆分别或一起在E.coil中表达,以用来确定每个亚基是否具有特异性功能.在基因表达的调节上,AGPase的调节方式属于转录水平调节.AGPase基因的mRNA转录物在花后15d达到最高水平,而此时淀粉的积累速率最快,表明基因的表达促进了淀粉的积累.在小麦子粒中,AGPase的含量和淀粉合成速率与AGPase的mRNA含量成正相关一.也曾有研究发现小麦开花后5d胚乳中有明显的mRNA积累, 并在开花后20~30d出现最高表达水平.相反,叶片中AGPase大小亚基mRNA水平显着差异与多肽水平的接近则说明了在叶片中转录后的调控起主要作用.3酶活性的调控3.1蔗糖合成酶研究表明,果糖和UDPG抑制SS的降解活性,而UDP抑制酶的合成活性,葡萄糖对合成和降解都有抑制作用;SS发挥活性需借助Mg,但zn,Hg2+,Cu2+,Fe,M,Co对酶活性均有抑制作用E3,387.SS活性通常在合成淀粉或者细胞壁的组织中最高,Stone等|4..指出,在淀粉合成过程中,各种酶的活性变化一般表现为单峰曲线,但酶活性的高低及其变化方式在基因型间有明显差132008.6作物杂志Crops异,依各地区生态条件和品种特性的不同而不同,此外,其活性大小也受到温度等外界环境条件的影响.3.2腺苷二磷酸葡萄糖焦磷酸化酶AGPase的活性调节包括变构调节和共价调节两种调控方式,对前者研究较为深入,而后者是通过铁氧还蛋白一硫氧还蛋白系统介导调节AGPase 活性,但其调控机理目前尚不清楚.Ghosh等¨认为AGPase是一个变构酶,主要表现为被3磷酸甘油酸(3-PGA)激活和被Pi抑制,当激活因子3-PGA与抑制因子Pi共同存在时, 将通过二者的比率来控制酶的活性,从而调控淀粉的合成.在谷类作物的子粒中也存在AGPase,但区别于叶绿体的AGPase,其活性对3一PGA和无机磷不敏感,这可能是由于单子叶植物的进化引起的,胞质内激活不敏感的AGPase是为适应胚乳发育的调控需要;可能是由于位于胞质的AGPase 可减少质体中ATP的产生和Pi循环J.Diego等也研究了对小麦胚乳和叶片中AGPase的调控特性,结果表明胚乳中的AGPase对3.PGA和磷酸的调节同样不敏感,而叶片中AGPase的活性能显着地被3-PGA激活和被磷酸抑制.AGPase除了可以被3-PGA变构激活外,二价阳离子Mg,Mn也可以对其变构激活.同时AGPase活性还受氧化还原势的修饰,还原力强时,二硫键断开,酶被激活.4研究展望植物生理学,分子生物学和基因工程的发展,对淀粉合成代谢途径的研究有了很大的突破,同时植物的高效遗传转化体系的建立,使得利用转基因的方法研究小麦子粒淀粉合成的途径成为可能,在此基础上可以通过研究控制胚乳淀粉合成的酶及基因来改变淀粉的产量和品质.ss是淀粉合成的关键酶,在小麦等作物产量调控方面具有重要的作用,可以借助分子生物学手段和反向遗传手段进行遗传转化,进一步深入研究小麦sS及其同工酶在植物体内的功能及作用机制,以寻求提高作物品质和产量的技术措施.AGPase具有表达的特异性,在不同组织,器官和不同植物中的结构和表达是不同的,可以通过基因手段将不同组织,器官,植物的AGPase基因14克隆转化来研究其性质.小麦中未见有AGPase突变体的报道,但Smidansky等将改良过的玉米AGPase大亚基基因2转入小麦并得到表达,发现每株子粒产量和整个植株的生物产量分别增加了38%和3l%.这一结果表明,转基因小麦由于增加了AGPase的活性,提高了ADPG的水平,进而增加了子粒的产量,因此有望通过提高AGPase的活性,增加小麦的产量.参考文献1黄琴,王志敏.禾谷类作物胚乳淀粉的生物合成.中国农业大学学报,1999,4(增刊):8~l52MaranaC,GarciaOF,CarboneroP.Differentexpressionoftwotypesof sucrosesynthase—encodinggenesinwheatinresponsetoanaembiosis, coldshockandlight.Gene,1990,88:167~1723卢合全,沈法富,刘凌霄等.植物蔗糖合成酶功能与分子生物学研究进展.中国农学通报,2005,21(7):34~374秦巧平,张上隆,谢鸣等.果实糖含量及成分调控的分子生物学研究进展.果树学报,2005,22(5):519~5255SturmA,TangGQ.Thesucrose—cleavingenzymesofplantsarecrucial fordevelopment,growthandcarbonpartitioning.TrendsinPlantSci (Reviews),1999,(4):401~4076张明方,李志凌.高等植物中与蔗糖代谢相关的酶.植物生理学通讯,2002,38(3):289~2957RuanYL,LlewellynDJ,FurbankRT.SuppressionofSUCroSesynthase geneexpressionrepressescottonfibercellinitiation,elongation,and seeddevelopment.PlantCell,2003,l5:952~9648李春燕,封超年,张影等.氮肥基追比对弱筋小麦宁麦9号子粒淀粉合成及相关酶活性的影响.中国农业科学,2005,38(6):1120一l1259王旭东,于振文,王东.钾对小麦旗叶蔗糖和子粒淀粉积累的影响.植物生态学报,2003,27(2):196~20110UmemotoT,NakamuraY,lahikuraN.Effectofgrainlocationonthe panicleonacfivitiesinvolvedinstarchsynthesisinriceendosperm. Phytochemistry,1994,36:843—8471l姜东,于振文,李永庚等.高产小麦强势和弱势子粒淀粉合成相关酶活性的变化.中国农业科学,2002,35(4):378—38312WangAY.KaoMH.Diferentiallyanddevelopmentallyregulated expressionofthreericesynthasegenes.PlantCellPhysiology,1999,4O(8):800~80713谢祝捷,姜东,曹卫星等.花后土壤水分状况对小麦子粒淀粉和蛋白质积累关键调控酶活性的影响.植物生理与分子生物学学报,2003,29(4):309~31614盛婧,郭文善,胡宏等.小麦淀粉合成关键酶活性及其与淀粉积累的关系.扬州大学学报(农业与生命科学版),2003,24(4):49~5315王月福,于振文,李尚霞等.小麦子粒灌浆过程中有关淀粉合成酶的活性及其效应.作物学报,2003,29(1):75~8l16KeelingPL,WoodJR,TysonRH.Starchbiosynthesisindeveloping wheatgrain.PlantPhysiology,1988,87:31l~31917NakamuraY,KukiK,ParkSY,eta1.Carbohydratemetabolismin thedevelopingendospermofricegrains.PlantCellPhysiology,1989,3O(6):833~83918JennerCF,AustJ.Effectsofexposureifwheatearstohightempera-作物杂志Crops2008.6 tureondrymatteraccumulationandcarbohydratemetabolisminthe grainoftwocuhivarsimmediateresponses.AustralianJournalofPlant Physio1.1991.18:165—177l9梁建生,曹显祖,徐生等.水稻子粒库强与其淀粉积累间关系的研究.作物学报,1994,20(6):686~69120熊瑛,李友军,郭天财.小麦淀粉合成相关酶的研究现状.河南科技大学学报(农学版),2004,24(2):6—921PreissJ,BallK,Smith—WhiteB,eta1.Starchbiosynthesisanditsreg—ulation.BiochemicalSocietyTransactions.1991,19:539~54722OkitaTW.Isthereanalternativepathwayforstarchsynthesis.Plant Physiology,1992,100:560—56423MoriguchiT.YamakiS.Purificationandcharaeterizationofsucrose synthasefromachfruit.PlantCellPhysiol,1988,29:1361~136624韩立敏,J{占之.植物水溶性蔗糖合成酶生物信息学分析初探. 植物生理学通氓,2006,42(5):939~94425SturmA.TangGQ.Thesucrose—cleavingenzymesofplantsarecruel? alfordevelopment.growthandcarbonpartitioning.TrendsinPlantSci(Reviews),1999,(4):401~40726HaouazineTN,TymowskalZ,TakV A,eta1.Characterizationoftwo membersoftheArabidopsisthalianagenefamily,codingforSUCrOSe syuthase.Gene,1997,197:239—25127ShaneCH.StevenCH.Proteasomeactivityandthepost-translational controlofsucrosesynthasestabilityinmaizeleaves.PlantPhysiology andBiochemistry2004,42:197~20828AkireK.Analysisofsucrosesynthasegenesincitrussuggestsdiffer—entrolesandphylogeneticrelationships.JournalofExperimentalBot- any,2002,53:61—7129AnisworthC.TarvisM.ClarkJ.IsolationandanalysisofcDNAclone encodingthesmallsubunitofADP?gluc0spyrephosphorylasefrom wheat.PlantMolecularBiology,1993,23:23—333O高振宇,黄大年.植物中合成蔗糖和淀粉的关键酶I.腺苷二磷酸-葡萄糖焦磷酸化酶.生命的化学,1998,18(3):26~28parisonofproteinofADP—glucosePY- rophorylasefromdiverse.JMolEvol,1992,34:449—46432MorellMK,RahmanS,AbrahamsSL,eta1.Thebiochemistryand molecularbiologyofstarchsynthesisincerea1.AustJPlantPhysiol, 1995,22:647~660ughlinMJ.FuYB.Adenosine5一diphosphate—Glucosepyrophosphorylasefrompotatotuber.PlantPhysiol,1995, 109:245~25134FuY,BallieoraMA,LeykamJF,eta1.Mechanismofreductiveacti. rationofpotatotuberADP-glucosepyrophosphorylase.JBiolChem. 1998.273:25045~2505235OliveMR.EllisRJ,SchuchWW.Isolationandnucleotidese, quenceofeDNAcloneencodingADP—glucosepyrophosphorylase polypeptidefromwheatleafandendosperm.PlantMolecularBiology, 1989,12:525~52836ReevesCD,KrishanHB,OldtaTW.Geneexpressionindevelop? mentwheatendosperm.PlantPhysiol,1986,82:34—4O37LeviC,PreissJ.RegulationpropertiesoftheADP—Glucosepyroplos- phorylaseoftheblue-greenSynechocccus.PlantPhysiol,1976,58: 753—75638EllingL.Effectofmetalionsonsucrosesynthasefromricegrains—a studyonenzymeinhibitionandenzymetopography.Glycobiology, 1995,52:201~20639MccollumTG.HuberDJ.CantlifeDJ.Solublesugaraccumulation andactivityofrelatedenzymesduringmuskmelonfruitdevelopment.JAmerSocHortSci.1988.1l3:399~40340StoneDJ,NicolasME.Varietaldifferenceinnaturalproteincomposi?tionofwheat.AustJPlantPhysiol,1996,23:727—73741GhoshHP,PreissJ.AdenosineDiphosphateGlucosePyrophosphory-lase.Aregulatoryenzymeinthebiosynthesisofstarchinspinachleafchloroplasts.JBiolChem,1966,241:449l一450442BallicraMA,LaughlinMJ,FuYB,eta1.Mechenismofreductive activationofpotatotuberADP—glucosepyrophosphorylase.JBiolche.1998,273:25045—2505243DenyerK,DunlapF,ThorbjornsenT,eta1.ThemaorformofADP—glucosepyrophosphorylaseinmaizeendospermisextra?plastidiai.PlantPhysiol,1996,112:779~78544包劲松,夏英武.水稻淀粉合成的的分子生物学研究新进展.植物学通报,1999,16(4):352~35845DiegoF,Gomez—Gasati,AlbeaoA,eta1.ADP?glucosepyrophospho—rylasefromwheatendosperm.Purificationandcharacterizationofan enzymewithnovelregulatoryproperties.Planta,2002,214:428~43446SmidanskyED,ClancyM,MeyerFD,eta1.EnhancedADP—glucose pyrophosphorylaseactivityinwheatendospermincreasesseedyield. PrecNatlAcadsciUSA,2002,99(3):1724~1729 RecentAdvancesonWheatSucroseSynthaseandADP-glucosePyr0ph0sph0rylaseJiangLina,LiDongfen,LiGhunxi,ShaoYun(CollegeofLifeSciences,HenanNormalUniversity,Xinxiang453007,Henan,China) AbstractStarchisanessentialconstituentofwheatendospermandthusisimportantforquality andyield.Su—erosesynthase(SS)andADP-glucosepyrophosphorylase(AGPase)playanimportantrolein plantstarchsynthe-sis.StudiesORtheseenzymesarebrieflyreviewedinthispaper,includingtheorientation,the biologicalfunctions,theanalysesbasedonmolecularbiology,andtheregulationofenzymaticactivitiesetc. KeywordsWheat;Sucrosesynthase;ADP-glucosepyrophosphorylase15。
水稻中两个二磷酸腺苷葡萄糖焦磷酸化酶基因及其编码蛋白的工程应用[发明专利]
![水稻中两个二磷酸腺苷葡萄糖焦磷酸化酶基因及其编码蛋白的工程应用[发明专利]](https://img.taocdn.com/s3/m/0e0c14ea0740be1e650e9af0.png)
专利名称:水稻中两个二磷酸腺苷葡萄糖焦磷酸化酶基因及其编码蛋白的工程应用
专利类型:发明专利
发明人:徐国华,顾冕,孟齐,张雯琦
申请号:CN202010937424.6
申请日:20200908
公开号:CN112048489A
公开日:
20201208
专利内容由知识产权出版社提供
摘要:本发明公开了水稻中两个二磷酸腺苷葡萄糖焦磷酸化酶基因及其编码蛋白的工程应用。
水稻二磷酸腺苷葡萄糖焦磷酸化酶基因OsAGPL1和/或OsAGPS1在调控水稻对磷素的吸收及利用中的应用。
一种促进水稻对磷素吸收和利用的方法,突变、沉默或抑制水稻中OsAGPL1和/或OsAGPS1的表达,以促进水稻对磷素吸收和利用。
突变OsAGPL1和OsAGPS1单/双基因,有利于提高水稻在磷供应充足条件下对磷素吸收及无机磷在水稻体内的积累。
申请人:南京农业大学
地址:211225 江苏省南京市溧水区白马镇国家农业科技园南京农业大学基地
国籍:CN
代理机构:南京天华专利代理有限责任公司
更多信息请下载全文后查看。
4.08级生化复习题-糖代谢参考答案

糖代谢名词解释:1.糖异生:非糖物质(如丙酮酸乳酸甘油生糖氨基酸等)转变为葡萄糖的过程。
2.Q酶:Q酶是参与支链淀粉合成的酶。
功能是在直链淀粉分子上催化合成(α-1, 6)糖苷键,形成支链淀粉。
3.乳酸循环乳:酸循环是指肌肉缺氧时产生大量乳酸,大部分经血液运到肝脏,通过糖异生作用肝糖原或葡萄糖补充血糖,血糖可再被肌肉利用,这样形成的循环称乳酸循环。
4.发酵:厌氧有机体把糖酵解生成NADH中的氢交给丙酮酸脱羧后的产物乙醛,使之生成乙醇的过程称之为酒精发酵。
如果将氢交给病酮酸丙生成乳酸则叫乳酸发酵。
5.变构调节:变构调节是指某些调节物能与酶的调节部位结合使酶分子的构象发生改变,从而改变酶的活性,称酶的变构调节。
6.糖酵解途径:糖酵解途径指糖原或葡萄糖分子分解至生成丙酮酸的阶段,是体内糖代谢最主要途径。
7.糖的有氧氧化:糖的有氧氧化指葡萄糖或糖原在有氧条件下氧化成水和二氧化碳的过程。
是糖氧化的主要方式。
8.肝糖原分解:肝糖原分解指肝糖原分解为葡萄糖的过程。
9.磷酸戊糖途径:磷酸戊糖途径指机体某些组织(如肝、脂肪组织等)以6-磷酸葡萄糖为起始物在6-磷酸葡萄糖脱氢酶催化下形成6-磷酸葡萄糖酸进而代谢生成磷酸戊糖为中间代谢物的过程,又称为磷酸已糖旁路。
10.底物水平磷酸化(substrate phosphorlation):ADP或某些其它的核苷-5′—二磷酸的磷酸化是通过来自一个非核苷酸底物的磷酰基的转移实现的。
这种磷酸化与电子的转递链无关。
11.柠檬酸循环(citric acid cycle):也称为三羧酸循环(TAC),Krebs循环。
是用于乙酰CoA中的乙酰基氧化成CO2的酶促反应的循环系统,该循环的第一步是由乙酰CoA经草酰乙酸缩合形成柠檬酸。
12.回补反应(anaplerotic reaction):酶催化的,补充柠檬酸循环中间代谢物供给的反应,例如由丙酮酸羧化酶生成草酰乙酸的反应。
13.乙醛酸循环(glyoxylate cycle):是某些植物,细菌和酵母中柠檬酸循环的修改形式,通过该循环可以收乙乙酰CoA经草酰乙酸净生成葡萄糖。
一个新的玉米Bt2基因突变体的遗传分析和分子鉴定

作物学报 ACTA AGRONOMICA SINICA 2022, 48(3): 572−579/ ISSN 0496-3490; CN 11-1809/S; CODEN TSHPA9E-mail: zwxb301@本研究由国家自然科学基金项目(U1804235, 31771800)和河南农业大学科技创新基金项目(KJCX2020A04)资助。
This study was supported by the National Natural Science Foundation of China (U1804235, 31771800) and the Science and Technology Innovation Fund of Henan Agricultural University (KJCX2020A04).*通信作者(Corresponding author): 陈洪宇, E-mail: chenhongyu@第一作者联系方式: E-mail: 158********@Received (收稿日期): 2021-01-19; Accepted (接受日期): 2021-06-16; Published online (网络出版日期): 2021-07-19. URL: https:///kcms/detail/11.1809.S.20210716.1506.004.htmlDOI: 10.3724/SP.J.1006.2022.13005一个新的玉米Bt2基因突变体的遗传分析和分子鉴定徐宁坤 李 冰 陈晓艳 魏亚康 刘子龙 薛永康 陈洪宇* 王桂凤河南农业大学农学院 / 省部共建小麦玉米作物学国家重点实验室, 河南郑州 450002摘 要: 玉米籽粒发育调控机制的研究对于玉米产量与品质性状的遗传改良十分重要。
本研究鉴定了一个新的转座子插入的籽粒皱缩突变体5601Q , 遗传分析表明其籽粒缺陷稳定遗传且为单基因隐性突变。
马铃薯腺苷二磷酸葡萄糖焦磷酸化酶基因克隆

马铃薯腺苷二磷酸葡萄糖焦磷酸化酶基因克隆马铃薯腺苷二磷酸葡萄糖焦磷酸化酶基因克隆马铃薯(Solanum tuberosum)作为一种重要的农作物,在全球范围内广泛种植。
马铃薯的主要产物为块茎,块茎中富含淀粉,是人类重要的食物来源之一。
而马铃薯腺苷二磷酸葡萄糖焦磷酸化酶基因(ADPG pyrophosphorylase)在马铃薯的淀粉代谢中起着重要的调控作用。
ADPG焦磷酸化酶是淀粉代谢途径中的关键酶之一,它通过将葡萄糖-1-磷酸(G1P)和腺苷二磷酸葡萄糖(ADPG)反应生成葡萄糖-6-磷酸(G6P)和焦磷酸酯(PPi),从而驱动了淀粉的合成。
淀粉的合成在马铃薯块茎的形成中起着重要的作用,因此ADPG焦磷酸化酶基因对马铃薯产量和品质具有重要的影响。
为了进一步研究马铃薯中ADPG焦磷酸化酶基因的功能,科学家们决定进行其基因的克隆工作。
克隆是一种将基因从其存在的生物体中分离提取,并通过特定的实验方法进行扩增和处理的技术。
在ADPG焦磷酸化酶基因的克隆中,科学家们首先从马铃薯组织中提取了总RNA,并通过逆转录反应将其转化为cDNA。
随后,利用PCR技术,他们扩增了ADPG焦磷酸化酶基因的全长cDNA序列。
通过PCR扩增后,科学家们将ADPG焦磷酸化酶基因的cDNA序列克隆到适当的载体上,以便后续的功能研究。
在克隆过程中,选择适当的载体对于保持基因的完整性和稳定性非常重要。
科学家们选择了一种常用的质粒载体,并通过限制性内切酶切割将ADPG焦磷酸化酶基因的cDNA插入到质粒中。
在转化完成后,科学家们对转化后的大肠杆菌进行筛选和培养,以获得包含ADPG焦磷酸化酶基因的菌落。
最终,经过筛选和鉴定后,科学家们成功获得了拥有马铃薯ADPG焦磷酸化酶基因的重组大肠杆菌。
获得ADPG焦磷酸化酶基因的重组大肠杆菌后,科学家们进一步对其进行酶活性和调控机制的研究。
通过测定酶活性和淀粉合成速率等指标,科学家们发现ADPG焦磷酸化酶在马铃薯淀粉代谢中具有重要的催化作用。
腺苷二磷酸葡萄糖焦磷酸化酶研究进展

高 淀 粉含 量和 改 良农作 物 品质具 有 重要 意义 .
1 AGP s 位 与 功 能 ae定
1 1 AGP s . a e定 位
的 定 位 、 能 、 学特 性 、 子 生 物 学 已经 进 行 了较 为 深入 和 系统 的 综 述 . 功 酶 分 关 键 词 : 苷 二 磷 酸 葡 萄 糖 焦磷 酸 化 酶 ; 基 ; 录 ; 粉 合 成 腺 亚 转 淀
中图 分 类 号 : 5 2 ¥ 3 文 献 标 志码 : A
摘 要 : 苷二 磷 酸 葡 萄 糖 焦 磷 酸 化 酶 ( P gu o e y ( h s h r l e AG a e 腺 AD — lc s p o p o ya , P s )是 植 物 和 细 茵 中淀 粉 和 糖 原 合 pr ) s
成 的 限速 酶 , 酶 催 化 l 酸 葡 萄 糖 ( 一 ) 三磷 酸 腺 苷 ( TP 反 应 形 成 腺 苷 二 磷 酸 葡 萄 糖 ( P . 关 该 酶 该 一 磷 G 1P 与 A ) AD G) 有
底物 , 淀粉 生物 合成 过程 中的关键 酶 , 主要 依据 是 其
是: ①在 AGP s 缺 失 的 突 变 体 中 淀 粉 的 含 量 明 显 ae
下 降. 玉米 胚 乳 s rn e h u k n2和 b i l一 r t 2突 变 体 ( te 分 别 为 AG ae大小 亚 基基 因 的突变 体 ) 仅 含 5 ~ Ps , 1 的酶活性 , O 淀粉 含量仅 占野 生 型 玉米 的 2 ~ 5
在 光合 细胞 中 AGP s a e定 位 于 质体 内 , 作 用 所 的底 物 G—_ lP和 ATP存在 于质 体 中. 在植 物非 光 而 合器 官 中 AG ae定 位 始 终 是 一个 有 争 议 的 问 题. Ps 在 叶绿体 中 , ATP来 自于 光 合 作用 , 在 非 光 合 器 而 官质 体 中 , ATP是 AD ( P 二磷 酸腺 苷 ) ATP转运 蛋 / 白从 胞质 溶胶 中输入 的. 叶绿 体 中 , 一一 在 G 1P是 通 过 还 原 性戊 糖磷 酸 循环 中葡 糖磷 酸异 构 酶和 葡糖磷 酸
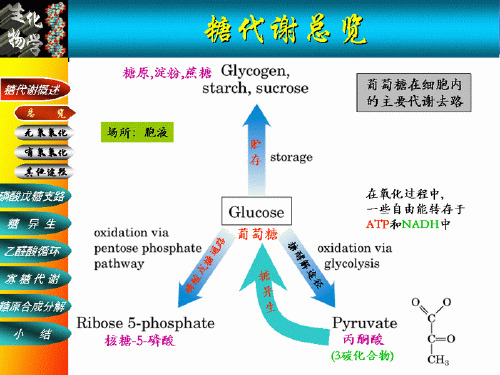
第七章 糖代谢
K2=3250
在植物光合组织中蔗糖磷酸合酶的活性较高,而非光合组 织中蔗糖合酶的活性较高。这是目前认为可能在光合组织中合 成蔗糖的主要途径。
(二)淀粉的合成:
存在于植物体内,尤其是谷类、豆类、薯类 作物的籽粒和贮藏组织都含丰富的淀粉。
淀粉合成中的糖基供体有ADPG、UDPG, 主要是ADPG。
合成分两阶段进行,先合成直链淀粉,然后 分支形成支链淀粉。
二、反应过程 反应可分为两个阶段: 第一阶段:氧化阶段,生成NADPH+H+和 CO2;由6-磷酸葡萄糖直接脱氢脱羧生 成磷酸戊糖; 第二阶段:非氧化阶段,一系列基团转 移反应;磷酸戊糖分子再经重排最终又 生成6-磷酸葡萄糖。
第一阶段:氧化阶段
1、脱氢反应:6-磷酸葡萄糖脱氢酶以NADP+ 为辅酶,催化6-磷酸葡萄糖脱氢生成6-磷酸 葡萄糖酸δ内酯,不可逆。
生物合成的供氢体
脂肪酸、胆固醇和类固醇化合物 的生物合成,均需要大量的NADPH。
0 R-CH2-C-R’
=
OH R-CH2-CH-R’ NADP+
R-CH2-CH2-R’
NADPH + H+
H R-C=C-R’
3、磷酸戊糖途径与疾病
神经精神病
(neuropsychiatric disorder)
第六节 糖的合成代谢
一、光合作用 二、糖异生途径 三、蔗糖和多糖的生物合成
一、光合作用
• 光合作用是糖合成代谢的主要途径。 • 绿色植物、光合细菌或藻类等将光能转变成化学 能的过程,即利用光能,由CO2和H2O合成糖类化 合物并释放出氧气的过程,称为光合作用。 • 光合作用的总反应式可表示如下: 光能 • n CO2 + n H2O (CH2O)n + n O2 叶绿体 糖类化合物
小麦腺苷二磷酸葡萄糖焦磷酸化酶同工酶基因型与酶活性及淀粉含量的关系

小麦腺苷二磷酸葡萄糖焦磷酸化酶同工酶基因型与酶活性及淀粉含量的关系岳向文;赵法茂;李天骄;王宪泽【期刊名称】《作物学报》【年(卷),期】2008(34)9【摘要】采用非变性聚丙烯酰胺凝胶电泳鉴定了我国60个代表性小麦品种的腺苷二磷酸葡萄糖焦磷酸化酶(AGP)同工酶基因型,并测定了AGP活性及总淀粉含量,以明确小麦籽粒AGP同工酶基因型组成及其与AGP活性和淀粉含量的关系.结果表明,AGP有AGPa、AGPb、AGPc和AGPd 4个等位基因位点,其出现频率分别为96.7%、80.0%、86.7%和16.7%:共检测到5种基因型,其中基因型AGPabc出现频率最高,为46.7%.不同基因型的品种间AGP活性和总淀粉含量差异显著(P<0.05).其中具有基因型AGPabcd的品种酶活性及总淀粉含量最高,表明小麦籽粒中不同AGP同工酶基因型对酶活性及淀粉含量有不同遗传效应.【总页数】6页(P1644-1649)【作者】岳向文;赵法茂;李天骄;王宪泽【作者单位】山东农业大学作物生物学国家重点实验室,山东泰安,271018;泰山学院生物系,山东泰安,271021;山东农业大学作物生物学国家重点实验室,山东泰安,271018;山东农业大学作物生物学国家重点实验室,山东泰安,271018【正文语种】中文【中图分类】S5【相关文献】1.小麦腺苷二磷酸葡萄糖焦磷酸化酶同工酶类型及时空表达分析 [J], 赵法茂;岳向文;杨红花;肖军;李运祥;王宪泽2.腺苷二磷酸葡萄糖焦磷酸化酶活性对小麦K、V、T型不育系育性及籽粒形成的影响 [J], 吴世文;高庆荣;孙哲;王茂婷;孙正娟;袁凯;于松3.小麦籽粒淀粉分支酶同工酶基因型与酶活性关系研究 [J], 赵法茂;毕建杰;李天骄;逄孝云;王宪泽4.转反义trxs基因小麦籽粒蛋白组分的巯基含量与α-淀粉酶活性的关系 [J], 周苏玫;尹钧;任江萍;张冉;李磊;郭红祥5.黄芪毛状根中尿苷二磷酸葡萄糖焦磷酸化酶活性与多糖含量关系的研究 [J], 吴晓俊;刘涤;胡之璧因版权原因,仅展示原文概要,查看原文内容请购买。
【国家自然科学基金】_腺苷二磷酸葡萄糖焦磷酸化酶_基金支持热词逐年推荐_【万方软件创新助手】_20140730

2012年 序号 1 2 3 4
科研热词 蔗糖合成酶 甘薯 多效唑 adpg焦磷酸化酶
推荐指数 1 1 1 1
2013年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14
科研热词 鳞茎 酶 籽粒灌浆 碳水化合物代谢 源--库转换 淀粉合成酶 水稻 抗旱性 干旱胁迫 小麦 反义trxs基因 单片段代换系 产量 东方百合
推荐指数 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
2010年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17
科研热词 推荐指数 淀粉 2 马铃薯块茎 1 酶活性 1 荧光定量rt-pcr 1 腺苷二磷酸葡萄糖焦磷酸化酶(agpase) 1 胞质型小亚基 1 玉米 1 淀粉含量 1 淀粉合成酶基因 1 淀粉合成酶 1 木薯 1 总rna提取 1 基因表达 1 同工酶位点 1 trizol法 1 chan法 1 agpase 1
推荐指数 1 1 1 1 1 1 1 1 1 1 1 1 1 1
2008年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18
科研热词 推荐指数 酶活性 3 淀粉 3 冬小麦 3 淀粉粒 2 高温 1 遗传转化 1 腺苷二磷酸葡萄糖焦磷酸化酶 1 番茄 1 淀粉合成关键酶 1 水稻 1 氮代谢 1 强筋 1 弱筋 1 弱光 1 土壤水分 1 品质 1 反义载体 1 乙烯 1
2009年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23
科研热词 非胚部位(胚乳) 酶活性 酶活力 菌株 荧光反应 腺苷二磷酸-葡萄糖焦磷酸化酶 紫外-可见分光光度法 籽粒发育 穗型 生理指标 玉米 淀粉 测定 水稻 根系 微胚乳玉米 强势粒 弱势粒 库强度 品质 化学信号 冬小麦 agpase
腺苷二磷酸葡萄糖焦磷酸化酶活性对小麦K、V、T型不育系育性及籽粒形成的影响

山东农业 大学 农学 院 /国家作 物生物 学重 点实验 室,山东 泰安 2 1 1 ; 7 0 8 泰安市 农业 科学研 究 院,山东泰 安 2 10 70 0
摘 要:为进 一步探 寻小 麦不育 系 的不 育机 制 和籽 粒不 饱满 的生理 机制 ,以冀 5 1 4 8核基 因为 遗传背 景,对 同核 异质
A g i u t r l c e c , l ’n 2 0 , i a rc lu a i n e Ta a 7 00 Ch n S 1
Ab ta t Cyo ls cmaese l CMS ie , s e il ( glp sr c : tpa mi l—tr e( i )l s ep cal K Ae i s幻f f, ( glp e tio a . n T ic m mo n y o c v) V Ae i sv nrc s ) a dT( rt u t — o i i
E mal x z — i b w@c iao r a.e.a : hn j un 1n t c
DoI 03 2 / PJ 1 0 .0 0 0 9 5 :1 .7 4 S ..0 62 1 .0 9
腺 苷 二磷 酸 葡 萄糖 焦磷 酸 化 酶 活 性 对 小 麦 K、V、T型 不 育系 育性 及 籽粒 形 成 的影 响
w u h— n ,GAO Si We Qi gRo g, UN h ,WANG Ma — i g ,S n . n ,S Z e , oT n UN Z e g J a h n .u n ,YUAN i,a d Ka n
Y U o S ng
Na o a y La o a o y o o o o y /A ̄o o i t n l Ke b r t r f Cr p Bi l g n my Co l g ,S a d g Ag i u t a l e h n on rc l e ur lUn v r i Ta ’n 27 01 , i e st y, l a 1 8 Ch n ; Ta ’ n Ac d my o i a ia a e f
6生物化学习题(答案)

6生物化学习题(答案)5 糖类分解代谢一、名词解释1、糖酵解途径:是在无氧条件下,葡萄糖进行分解,形成2分子丙酮酸并伴随着ATP生成的一系列反应。
2、柠檬酸循环:是用于乙酰CoA中的乙酰基氧化生成CO2的酶促反应的循环系统,该循环的第一步反应是由乙酰CoA和草酰乙酸缩合形成柠檬酸。
3、糖的有氧氧化:糖的有氧氧化指葡萄糖或糖原在有氧条件下氧化成水和二氧化碳的过程。
是糖氧化的主要方式。
4、磷酸戊糖途径:是指机体某些组织(如肝、脂肪组织等)种一个葡萄糖-6-磷酸经代谢产生NADPH和核糖-5-磷酸的途径。
该途径包括氧化和非氧化两个阶段,在氧化阶段,葡萄糖-6-磷酸转化为核酮糖-5-磷酸和CO2,并生成两分子的NADPH;在非氧化阶段,核酮糖-5-磷酸异构化生成核糖-5-磷酸或转化为酵解中的两个中间代谢物果糖-6-磷酸和甘油醛-3-磷酸。
5、发酵:厌氧有机体把糖酵解生成NADH中的氢交给丙酮酸脱羧后的产物乙醛,使之生成乙醇的过程称之为乙醇发酵。
如果将氢交给丙酮酸生成乳酸则叫乳酸发酵。
二、填空1、糖酵解过程中有3个不可逆的酶促反应,这些酶是磷酸果糖激酶、己糖激酶和丙酮酸激酶。
2、3-磷酸甘油醛脱氢酶酶催化的反应是EMP途径中的第一个氧化反应。
3、糖酵解中催化作用物水平磷酸化的两个酶是磷酸甘油酸激酶和丙酮酸激酶。
4、在糖酵解中提供高能磷酸基团,使ADP磷酸化成A TP的高能化合物是1,3-二磷酸甘油酸和PEP。
5、糖酵解在细胞的细胞质中进行,该途径是将葡萄糖转变为丙酮酸,同时生成ATP和NADH的一系列酶促反应。
6、丙酮酸还原为乳酸,反应中的NADH来自于3-磷酸甘油醛的氧化。
7、TCA循环的第一个产物是柠檬酸。
由柠檬酸合酶,异柠檬酸脱氢酶,和α-酮戊二酸脱氢酶所催化的反应是该循环的主要限速反应。
8、TCA循环中有二次脱羧反应,分别是由异柠檬酸脱氢酶和α-酮戊二酸脱氢酶催化。
脱去的CO2中的C原子分别来自于草酰乙酸中的C1和C4。
高能磷酸化合物英文缩写

高能磷酸化合物英文缩写高能磷酸化合物的生物化学意义高能磷酸化合物(High-Energy Phosphate Compounds)是一类含有高水平化学能的化合物,它们在细胞的能量代谢中发挥至关重要的作用。
这些化合物通过释放能量驱动各种重要的生物化学过程。
常见类型最常见的高能磷酸化合物包括:三磷酸腺苷(ATP)二磷酸腺苷(ADP)一磷酸腺苷(AMP)肌酸磷酸(Creatine Phosphate)焦磷酸盐(Pyrophosphate)腺苷三磷酸(ATP)ATP是细胞中最突出的高能磷酸化合物,被称为“能量货币”。
ATP由腺苷(一种核苷)和三个磷酸基团组成。
这些磷酸基团之间的磷酸酐键含有大量的能量,在水解过程中可以释放能量。
ATP在细胞能量代谢中的作用ATP在细胞中扮演着关键角色,它为几乎所有需要能量的细胞过程提供动力,包括:肌肉收缩主动运输合成代谢反应神经冲动的传导ATP的再生当ATP被水解释放能量时,它会变成ADP。
ADP可以通过以下方式再生为ATP:细胞呼吸:在细胞呼吸过程中,糖类和脂肪被分解,产生的能量用于通过氧化磷酸化合成ATP。
底物水平磷酸化:某些生化反应直接将磷酸基团转移到ADP上,形成ATP。
肌氨磷酸肌氨磷酸是一种高能磷酸化合物,主要存在于骨骼肌中。
它通过将磷酸基团转移到ADP上迅速再生ATP,从而支持快速而短暂的肌肉活动。
焦磷酸盐焦磷酸盐是一种高能磷酸化合物,在核酸代谢中扮演着重要角色。
它通过水解释放能量,为DNA和RNA的合成提供动力。
高能磷酸化合物的调节高能磷酸化合物的水平受到严格的调节,以确保细胞中的能量供应与需求相匹配。
调节机制包括:激素(如胰岛素和肾上腺素)神经信号氧气可用性结论高能磷酸化合物是细胞能量代谢的支柱。
它们提供能量,支持各种生物化学过程,对于维持细胞的正常功能至关重要。
理解这些化合物的性质和作用对于了解细胞生物学的复杂性至关重要。
腺苷二磷酸葡萄糖焦磷酸化酶活性测定试剂盒说明书

腺苷二磷酸葡萄糖焦磷酸化酶(ADP-glucose pyrophosphorylase,AGP)活性测定试剂盒说明书(货号:G0536F分光法48样)一、产品简介:ADPG焦磷酸化酶(AGP,EC2.7.7.27)是植物淀粉合成过程中起关键性调节作用的酶,催化l-磷酸葡萄糖(G-1-P)与三磷酸腺苷(ATP)反应形成淀粉合成的直接前体腺苷二磷酸葡萄糖(ADPG),在植物中,主要存在于贮藏器官和叶片中。
AGP催化的逆向反应生成G1P,在反应体系中添加的磷酸己糖变位酶和6-磷酸葡萄糖脱氢酶依次催化生成6-磷酸葡萄糖酸和NADPH,340nm下测定NADPH增加速率,即可计算AGP活性。
二、测试盒组成和配制:试剂名称规格保存要求备注提取液液体60mL×1瓶4℃保存试剂一粉体mg×1支-20℃保存临用前甩几下使粉体落入底部,再加1.1mL蒸馏水溶解,仍-20℃保存。
试剂二粉体mg×1支4℃保存临用前甩几下使粉体落入底部,再加1.1mL蒸馏水溶解。
仍4℃保存。
试剂三液体32mL×1瓶4℃保存试剂四粉体mg×1支-20℃保存临用前甩几下使粉体落入底部,再加2.1mL蒸馏水溶解,仍-20℃保存。
【注】:粉剂量在mg级别,使用前用手甩几次或者进行离心,打开直接加入要求的试剂即可。
三、所需的仪器和用品:可见分光光度计、1ml石英比色皿(光径1cm)、水浴锅、台式离心机、可调式移液器、研钵、冰和蒸馏水。
四、腺苷二磷酸葡萄糖焦磷酸化酶(AGP)活性测定:建议正式实验前选取2个样本做预测定,了解本批样品情况,熟悉实验流程,避免实验样本和试剂浪费!1、样本制备:①组织样本:称取约0.1g组织(水分充足的样本可取0.2g),加1mL提取液,进行冰浴匀浆,12000rpm,4℃离心10min,取上清,置冰上待测。
【注意】若样本颜色较深(如较深颜色的植物叶片),可引起起始值A1值较大如超过1.5,可在样本制备过程中增加除色素步骤:取约0.1g组织(水分充足的样本可取0.5g),加入80%乙醇冰浴匀浆,12000rpm,4℃离心10min,弃掉色素较深的上清液;以上除色素步骤重复2次。
糖异生

糖的异生作用
1, 高水平的A酶,而变构激活二磷酸果糖酯酶 2, Pi、AMP、ADP变构激活磷酸果糖激酶和丙酮酸 激酶,而变构抑制二磷酸果糖酯酶
3, ATP/ADP比值高时EMP途径关闭、糖异生打开 ; ATP/ADP比值低时, EMP途径打开,糖异生活 性降低
(一)直链淀粉的合成 4、蔗糖转化为淀粉
二. 淀粉的合成
在植物细胞中,淀粉合成的糖基大多来源于蔗 糖。在蔗糖合酶的催化下,蔗糖中的葡萄糖基转 移到ADP(UDP)上,形成ADPG,然后在淀粉 合酶的催化下,ADPG将其葡萄糖基转移到淀粉 的非还原端,使淀粉链延长。
(一)直链淀粉的合成 4、蔗糖转化为淀粉
2, 糖尿病人或切除胰岛的动物体内,从氨基酸转 化为糖的过程十分活跃
二、生物化学过程 1. 丙酮酸生成磷酸烯醇式丙酮酸
糖的异生作用
CO2 乙酰CoA +Mg2+ + ATP
丙酮酸
丙酮酸羧化酶 +生物素
草酰乙酸
丙酮酸羧化酶定位于线粒体
二、生物化学过程 1. 丙酮酸生成磷酸烯醇式丙酮酸
糖的异生作用
1、磷酸蔗糖合酶
一. 蔗糖的合成
G-1-P + UTP + H2O → UDPG + 2Pi
(UDPG焦磷酸化酶,焦磷酸酶)
该反应的自由能变化很小,反应是可逆的。但 由于细胞内的焦磷酸酯酶能及时将焦磷酸水解 成2分子磷酸,从而使反应向生成UDPG的方向 进行。
1、磷酸蔗糖合酶
一. 蔗糖的合成
UDPG + F-6-P → 磷酸蔗糖 + UDP
1,4→1,6转葡糖基酶(切割,连接)
(一)直链的合成
三. 糖原的合成
植物葡萄糖焦磷酸化酶

植物葡萄糖焦磷酸化酶
植物葡萄糖焦磷酸化酶是一种重要的酶类,它能够催化葡萄糖分子与ATP(三磷酸腺苷)经历葡萄糖激酶和葡萄糖-6-磷酸异构酶的作用后生成葡萄糖6-磷酸。
这个过程就是葡萄糖焦磷酸化作用。
植物葡萄糖焦磷酸化酶主要存在于植物中的叶片、根、种子、果实等组织中,其含量与光照强度、养分供应、外界环境等因素密切相关。
该酶在植物体内能够有效地参与光合作用和细胞代谢过程中的二磷酸腺苷(ADP)和ATP的转化,有助于维持植物体内生化代谢平衡和能量供应。
此外,植物葡萄糖焦磷酸化酶在植物的生长发育、抗逆应答和营养调控等方面也发挥着重要作用。
因此,研究植物葡萄糖焦磷酸化酶及其调控机制对于揭示植物的生长发育、代谢调节等方面的机理具有重要的理论和实践价值。
生化英文缩写介绍知识分享

生物化学常用英文缩写DNFB 2,4-二硝基氟苯DNS-Cl 丹磺酰氯FAD 黄素腺嘌呤二核苷酸IU 国际酶活力单位Vit 维他命TPP 硫胺素焦磷酸FH4 四氢叶酸AMP 腺苷一磷酸ADP 腺苷二磷酸ATP 腺苷三磷酸HA 透明质酸CS 硫酸软骨素KS 硫酸角质素HS 硫酸类肝素Hp 肝素PG 蛋白聚糖GPC 凝胶渗透层析HPGPC 高效凝胶渗透层析FA 不饱和脂肪酸PG 前列腺素LT 白三烯MDA 丙二醛TBA 硫代巴比妥酸SOD 超氧化物歧化酶GSHPX 谷胱甘肽过氧化物酶PAF 血小板活化因子PITC 苯异硫氰酸酯PTC 苯氨基硫甲酰PTH 苯异内酰硫脲5-FU 5-氟尿嘧啶5-HT 5-羟色胺6-MP 6-巯基嘌呤A/G 白球蛋白比率ACD 酸性枸橼酸葡萄糖ACP 酸性磷酸酶ACTH 促肾上腺皮质激素ADH 抗利尿激素ADP 二磷酸腺苷AFP 甲(种)胎(儿)蛋白AHF 抗血友病因子AKP 碱性磷酸酶ALL 急性淋巴细胞性白血病ALS 抗淋巴细胞血清ALT(GPT) 丙氨酸转氨酶AMA 抗线粒体抗体AMI 急性心肌梗塞AML 急性粒细胞性白血病AMP 一磷酸腺苷ANA 抗核抗体APC 复方阿司匹林AST(GOT) 天冬氨酸转氨酶ATN 急性肾小管坏死ATP 三磷酸腺苷BCG 卡介苗BCNU 卡氮芥BMR 基础代谢率BP 血压BSA (1)牛血清白蛋白;(2)体表面积BSP 磺溴酞钠BT 出血时间BUN 血尿素氮BV 血容量C3 补体3cAMP 环磷酸腺苷CAPD 不卧床的持续性腹膜透析CB-1348 苯丁酸氮芥CBG 皮质类固醇结合球蛋白CCNU 环己亚硝脲CCU 冠心病监护病室CEA 癌胚抗原cGMP 环磷鸟苷CIC 循环免疫复合物CIEP 对流免疫电泳CLL 慢性淋巴细胞性白血病CML 慢性粒细胞性白血病CMV 巨细胞病毒CNS 中枢神经系统CO2cp 二氧化碳结合力CoA 辅酶ACO 心输出量CPBA 竞争性蛋白结合分析CPC 临床病理讨论会CPK 肌酸磷酸激酶cpm 每分钟计数CSF 脑脊髓液CT (1)电算机体层摄影;(2)血凝时间CVP 中心静脉压DIC 播散性血管内凝血DMSA 二巯基丁二酸DNase 脱氧核糖核酸酶DNA 脱氧核糖核酸DNCB 二硝基氯苯DOCA 醋酸脱氧皮质酮dpm 每分钟衰变数DSA 数字减影血管造影DSS 登革休克综合征DTPA 二乙三胺五乙酸ECG;EKG 心电图EDTA 乙二胺四乙酸EEG 脑电图EHF 流行性出血热EIA 免疫酶法ELISA 酶联免疫吸附试验EMCP 肌电图ENT 耳鼻咽喉ERCP 逆行胰胆管造影ESR 红细胞沉降率FSH-RH 促卵泡素释放激素FSH 促卵泡素FT4I 游离甲状腺素指数FT4 游离甲状腺素GABA α-氨基丁酸GH 生长激素GTT 葡萄糖耐量试验HAI;HI 血凝抑制试验HA V 甲型肝炎病毒HBcAg 乙型肝炎核心抗原HBeAg 乙型肝炎e抗原HBsAg 乙型肝炎表面抗原HBV 乙型肝炎病毒Hb 血红蛋白HCG 人绒毛膜促性腺激素HCV 丙型肝炎病毒HDL 高密度脂蛋白HDV 丁型肝炎病毒HEV 戊型肝炎病毒HIV 人粪免疫缺陷病毒HLA 人白细胞抗原系统HP 高倍视野ICU 危重症病人监护病室IFA 免疫萤光法Ig 免疫球蛋白ITP 特发性血小板减少性紫癜IU 国际单位IVU 静脉尿路造影JBE 日本乙型脑炎keV 千电子伏LATS 长效甲状腺刺激素LD50 半致死量LDH 乳酸脱氢酶LDL 低密度脂蛋白LH-RH 黄体生成素释放激素LH 黄体生成素LP 低倍视野MAA 巨聚白蛋白MCHG 平均红细胞血红蛋白浓度MCV 平均红细胞体积MDP 亚甲基二磷酸MRI(NMR) 磁共振(核磁共振)成像MTX 氨甲蝶呤NHL 非何杰金淋巴瘤NPN 非蛋白氮OT 旧结核菌素PaCO2 动脉血二氧化碳分压PAGE 聚丙烯酰胺凝胶电泳PaO2 动脉血氧分压PAS 对氨基水杨酸PBI 蛋白结石碘PBS 磷酸盐缓冲液PB 磷酸盐缓冲剂PCR 聚合酶链反应PG 前列腺素PHA 植物血凝素:被动血凝试验pH 酸碱度(氢离子浓度负指数)PPD 结核菌素纯蛋白衍生物PSP 酚磺酞(酚红)PTC 经皮肝穿刺胆道造影PTH 甲状旁腺激素PT 凝血酶原时间rbc 红细胞RBC 红细胞计数RES 网状内皮系统RF 类风湿因子RIA 放射免疫分析RNA 核糖核酸rpm 每分钟转速(现改用r/min)RV 轮状病毒SaO2 氧饱和度SBE 亚急性细菌性心内膜炎SLE 系统性红斑狼疮SPECT 单光子发射计算机断层图像检查T3 三碘甲状腺原氨酸T4 甲状腺素TBG 甲状腺素结合球蛋白TG 甘油三酯Tg 甲状腺球蛋白TRH 促甲状腺激素释放激素TSH 促甲状腺激素TTT 麝香草酚浊度试验U 单位VLDL 极低密度脂蛋白Wbc 白细胞WBC 白细胞计数WHO 世界卫生组织.P.T.T.明矾沉淀破伤风类毒素A.T.S.破伤风血清A.T.旧结核菌素A/G ratio 白蛋白-球蛋白比率AA 氨基酸;变应性哮喘;AAA 美国变态反应学会AAbs 自身抗体AACR 美国癌症研究会AAE 急性变应性脑炎AAF α-干扰素激活的因子AAI 美国免疫学家协会AAS 抗炭疽血清AAV 腺相关病毒ab.抗体Abn 异常的ABO ABO分型法(血型)ABO system 人ABO血型系统ABS E 绝对误差ABS 抗原结合部位abs.alc.无水酒精Abstr 摘要Ac 抗补体AC 吸收系数ACA 自动皮肤过敏反应ACC 事故的;含抗体的细胞ACH 肾上腺皮质激素ACIF 抗补体免疫荧光AcOH 乙酸ACP 替代补体途径ACR 急性细胞排斥反应,获得性细胞性抵抗力ACTH 促肾上腺皮质激素ACTP 促肾上腺皮质激素多肽ACV 变应性皮肤血管炎AD virus 腺病毒AD 抗原决定簇(基)Ad 腺嘌呤AD50 50%激活剂量ADCC 抗体依赖性补体介导的细胞毒作用ADH 抗利尿激素ADIF 增效直接免疫荧光(试验)ADNAA 抗脱氧核糖核酸抗体ADP 腺苷二磷酸ADR DNA复制的激活酶ADR 药物不良反应AdR 脱氧核糖腺苷ADRIS 抗犬红细胞免疫血清ADRV 成人腹泻性轮状病毒ADT 琼脂凝胶扩散试验AE 绝对误差;琼脂糖凝胶电泳AED 急救容许剂量AEF 同种效应因子a-ELISA 放大酶联免疫吸附试验AET-STBC 氨乙基异硫脲处理过的绵羊红细胞AEX 阴离子交换AF 激活功能AFB 抗酸杆菌AFC 抗体形成细胞aFGF 酸性成纤维细胞生长因子AFP 甲胎蛋白Ag 抗原AG 琼脂糖;分析级别AGA 抗免疫球蛋白G自身抗体AGAT 抗球蛋白抗体试验AGD 琼脂凝胶扩散AGD(AD) 琼脂凝胶扩散Agg 凝集反应AGGS 抗气性坏疽血清AGMKC 非洲绿猴肾细胞AgR 抗原受体AH 抗组胺;急性肝炎AHC 急性出血性结膜炎AHDU 血吸附单位AHG 凝聚的人丙种球蛋白AI 自身抑制的AIC 自身免疫复合物AID 急性传染病AIDS 艾滋病aim.E.绝对误差AIP 过继性免疫预防AK cell 异常杀伤细胞ak 明显亲和常数ALA 抗淋巴细胞抗体alb 白蛋白alloAb 同种抗体alloAb 同种抗原ALT 丙氨酸转移酶amp 安瓿Amp 氨苄青霉素AMR 平均最小需要量Anti-HA V 抗甲型肝炎病毒的抗体Anti-HBc 抗乙型肝炎核心抗原的抗体Anti-HBc 抗乙型肝炎核心抗原Anti-HBs 抗乙型肝炎表面抗原的抗体Anti-Id 抗独特型抗体AP 过敏性紫癜APB 成人外周血APC 补体替代途径App 阑尾炎Appx 附录AR 过敏性反应;急性排斥反应;抗原比率ARD 自身免疫性风湿病ARDS 急性呼吸窘迫综合征AS 抗血清ASC 抗体分泌细胞ASLE 急性全身性红斑狼疮ASP 阿司匹林ATC 活化T细胞;人工靶细胞ATG 抗破伤风球蛋白ATP 三磷酸腺苷ATS 抗破伤风血清AU 吸收单位;抗体单位;抗毒素单位AU-PAGE 酸性尿素聚丙烯酰氨凝胶电泳autop 尸检A VP 抗病毒蛋白B 返回顶部B cellB淋巴细胞b.i.d 每日两次bac. 细菌学BAF B细胞活化因子Bas 嗜碱性粒细胞BBB 血脑屏障BBS 巴比妥缓冲盐水BBT 基础体温BC 血细胞计数BCG 卡介苗;牛丙种球蛋白BCGF B细胞生长因子BCG-PSN 卡介苗多糖核酸组分BCG 卡介苗BCL B细胞系BCR B细胞抗原受体BCR 无菌室BCSF B细胞刺激因子bFGF 碱性成纤维细胞生长因子BFP 生物假阳性(试验)B-G 百日咳杆菌BG 血型BI 加强免疫;结合指数;牛胰岛素biAb 双特异性抗体BIL 胆红素试验BM 基础培养基Bm 记忆性B淋巴细胞b-MAb 生物素化单克隆抗体Bmax 最大结合能力BMC 骨髓细胞BMD 骨盐密度BMI 体重指数BMR 基础代谢率BMT 骨髓移植BOD 生化需氧量BRM 生物反应调节剂BS 缓冲盐水BSA 牛血清白蛋白BSF B细胞刺激因子BSR 血细胞沉降率BU 生物学单位BWt 体重C 返回顶部顶部C系数;补体c,C 浓度C.F.T.补体结合试验c.p.m.次/分(每分钟次数)C.P.化学纯[净]Ca 癌CA 胶原蛋白CAH 慢性活动性肝炎CAH-B 慢性活动性乙型肝炎cal 卡Cam 氯霉素CAMS 中国医学科学院Cap 辣椒素cap 毛细血管CAR 趋化因子受体CAS 中国科学院CB 脐带血;B组柯萨奇病毒cc 立方厘米CCA unit 鸡(红)细胞凝集反应单位CCCCM 中国微生物菌种保藏管理委员会CCE 对流离心洗脱法CCM 完全培养基CCP 补体经典途径CD 临界剂量CD50 半数有效量CDC 疾病预防控制中心;补体依赖性细胞毒作用CDP 传染病防治计划CE 鸡胚;克隆效率;粗提取物CEF 集落促进因子CER 阳离子交换树脂CF 补体结合;细胞毒因子CFA 细胞荧光分析CFP(CTEP) 对流电泳(对流免疫电泳)CFR 病死率CFT 补体结合试验CFU-C 培养物中生长细胞的集落形成单位CFU-GM 粒细胞和巨嗜细胞的集落形成单位CF 补体结合CGL 慢性粒细胞白血病cGMP 环鸟苷酸CGN 慢性肾小球性肾炎Ch 慢性的;儿童CH50 50%补体溶血单位CHB 慢性乙型肝炎CHGP 中国人类基因组计划CHO 中国仓鼠卵巢Chr 染色体CHS 接触性超敏反应ChT 霍乱毒素Cht 趋化性CI 细胞毒指数;结合指数;抑制细胞生长指数CIE 对流免疫电泳;交叉免疫电泳CIEIA 竞争抑制酶免疫分析法CIEP 对流免疫电泳CIF 集落抑制因子CIS 中枢免疫系统CK 趋化因子;细胞因子CM 培养基;条件培养基CMA 中华医学会CMC 细胞介导的细胞毒作用CMIR 细胞介导免疫应答CMs 氯霉素敏感的Co Q 辅酶QCoA 辅酶ACOA 协同凝集反应COD 化学需氧量Col 秋水仙素COV 判定值CP 卡值;经典途径;氯霉素CPA 细胞增殖试验CPH 慢性迁延性肝炎CPR 瞳孔反应cps 每秒计数CPV 短小棒状杆菌菌苗CR 补体受体;条件反射CRIE 交叉放射免疫电泳Cs 培养物上清液CS 小牛血清;硫酸软骨素;接触性过敏CSF 脑脊液CSH 慢性重症肝炎CSM 脑脊髓膜炎CSN 对照上清液CSR 概略存活率CT 小牛胸腺;霍乱毒素CTE 小牛胸腺提取物CTX 细胞毒素CU 细胞毒单位CV 变异系数;柯萨奇病毒CVF 中心视野D 返回顶部D.L.M.最小致死量D.T.N.标准白喉毒素D/W 葡萄糖水溶液DASP method 双抗体固相法DAT 白喉抗毒素(拉)dB 分贝DB 稀释缓冲液DB-CT 双盲对比试验DC 扩散常数DD 双向扩散DDW 双蒸水DE 丧失视力Deg 度,程度DEM 直接电镜检验法DEN 登革病毒DEX 地塞米松DF 登革热DG 暗视野DH 迟发性超敏反应DHF 登革出血热DI 白喉;扩散指数DID 双向免疫扩散DIF 直接免疫电泳法DIF 直接免疫荧光DIG-ELISA 凝胶扩散-酶联免疫吸附试验Dip 白喉DLT 双标记技术DM 糖尿病DMC 双标记细胞DNA 脱氧核糖核酸DNase 脱氧核糖核酸酶DNA 脱氧核糖核酸DNOA 攻击死亡数DOA 到达时已死DOD 死亡日期DP 分辨力;双阳性dps 每秒种衰变数DPT 白喉、百日咳、破伤风DR 变性反应DST 脱敏试验;脱敏时间DST 脱敏试验DT 白喉毒素、白喉类毒素、白喉和破伤风类毒素DTH 迟发性超敏反应DTN 白喉标准毒素DTT 白喉破伤风类毒素DTx 白喉类毒素DW 蒸馏水E.D.有效量E 返回顶部E:T效靶比率Ea rosette 活性红细胞花环EAA 必须氨基酸EAC rosette 红细胞-抗体-补体花环EAC 红细胞抗体补体EAHF 湿疹、哮喘、枯草热EBV EB病毒;埃-巴二氏病毒EC50 有效浓度ECBS 生物制品标准化专家委员会ECD 电化学检测ED 有效剂量ED50 50%有效剂量ED50 半数有效量EEG 脑电图EF 外源因子EFA 变态反应增强因子EFT 红细胞玫瑰花结形成试验EG 卵清蛋白EHF 流行性出血热病毒;埃博拉出血热EHFV 流行性出血热病毒EI 酶联免疫吸附试验指数EIA 酶免疫试验EID 电免疫扩散ELISA 酶结合免疫吸附测定;酶标记免疫吸附分析法ELISA 酶联免疫吸附试验ELONP 酶标记寡核苷酸探针EMIT 酶放大免疫测定技术EP 流行性腮腺炎EP 内原性热原EP 英国专利;红细胞生成素EPI 扩大免疫接种规划EPI 扩大计划免疫ER 早期反应ERFT E玫瑰花法形成试验ERIA 电放射免疫分析Es 绵羊红细胞ES 内分泌系统ESR 血沉ETH 早期超敏反应EU 酶联免疫吸附试验单位EVC 霍乱弧菌F 返回顶部f.c. 最后浓度f.p. 冰点,凝固点F.P. 法国专利F/P 荧光染料与蛋白质之比FA 荧光抗体FAT 荧光抗体技术FB 成纤维细胞FBC 全血细胞计数FCA 弗氏完全佐剂FCS 胎牛血清FD 冷冻干燥FDA Food and DrugAdministration 美国药品食品管理局FD50 半数致死量FI 荧光强度FIA 荧光免疫测定法;弗氏不完全佐剂FIV 福尔马林灭活疫苗flu 流行性感冒FMD 口蹄疫Fn 纤维蛋白FPLC 快速蛋白质液相层析FPSE 甲醛丙酮醛固定的绵羊红细胞fr. 频率FS-ELISA 荧光夹心酶联免疫吸附试验FT4I 游离甲状腺素指数FU 荧光单位F-VC 流量容积曲线G 返回顶部GAP中药材生产质量管理规范GC 气相色谱法GCP Good ClinicPractice 药物临床实验管理规范G-CSF 粒细胞集落刺激因子GD 凝胶扩散GEF 凝胶等电点聚焦GF 凝胶过滤GFC 凝胶过滤色谱法GIT 生长抑制试验Glc 葡萄糖GLP GoodLaboratory Practice药物非临床研究质量管理规范Gm 庆大霉素GM 生长培养基GM-CFU 粒细胞-巨噬细胞集落形成单位GM-CSF 粒细胞-巨噬细胞集落刺激因子GMP 优良生产操作规程GMT 几何平均滴度GN 革兰氏阴性GNB 革兰氏阴性细菌GP 革兰氏阳性GPB 革兰氏阳性细菌GPC 气相分配色谱法GPP Good Processing Practice forper-manufacturing traditional Chinese medicines中药饮片生产质量管理规范GPT 谷丙转氨酶GR 肉牙肿GS 人参造甙GSC 气固色谱法GSP Good SupplyPractice 药品经营质量管理规范GTR 粒细胞周转率GTT 葡萄糖耐量试验Gua 鸟嘌呤GV 龙胆紫GVHR 移植物抗宿主反应H 返回顶部HA甲型肝炎HA 血细胞凝集[作用]HAE 血细胞比容HAI 血球凝集试验HB 乙型肝炎HBAg 乙型肝炎抗原HbcAb 乙型肝炎(病毒)核心抗体HBcAb 乙型肝炎核心抗体HbcAg 乙型肝炎(病毒)核心抗原HBcAg 乙型肝炎核心抗原HBD 健康供血者HBeAb 乙型肝炎e抗体HbeAg 乙型肝炎(病毒)e抗原HBeAg 乙型肝炎e抗原HBIg 乙型肝炎免疫球蛋白HBIG 乙型肝炎免疫球蛋白HbsAb 乙型肝炎(病毒)表面抗体HbsAg 乙型肝炎(病毒)表面抗原HBsAg 乙型肝炎表面抗原HBV 乙型肝炎病毒HC 健康对照;丙型肝炎HCC 肝癌Hct 血细胞比容HCV 丙型肝炎病毒HD 血液透析;丁型肝炎;高密度;高剂量HD50 半数溶血量HDF 血液滤过HDI 宿主防御指数HDN 新生儿溶血性疾病HDV 丁型肝炎病毒HE 戊型肝炎HEK 人胚肾HEL 人胚肺Hep 肝素HEV 戊型肝炎病毒HF 辅助因子HFRS 合并肾综合征的出血热HIg 人免疫球蛋白Hly 溶血素HMW 高分子量HOT 人型旧结核菌素HPAC 高亲和层析HPAEC 高效阴离子交换液相色谱(法)HPEC 高效电泳色谱法HPF 高倍视野HPLC 高压液相色谱(法)HPV 人乳头(状)瘤病毒HR 高应答者HRCV 人呼吸冠状病毒HRF 组胺释放因子HRIG 人狂犬病免疫球蛋白HRP 辣根过氧化物酶HRS 马血清HRV 人轮状病毒HS 人血清HSA 人血清白蛋白HSCT 造血干细胞移植HSE 人脾脏提取物HSV-Ⅰ单纯疱疹病毒Ⅰ型HSV-Ⅱ单纯疱疹病毒Ⅱ型HU 溶血单位HuIFN 人干扰素HuIFN-α 人α干扰素HuIFN-β 人β干扰素HuRBC 人红细胞HV 汉坦病毒属HWS 人全血清HZV 带状疱疹病毒HZ 带状疱疹Hφ 疏水性I 返回顶部i.c. 皮内注射i.m. 肌内(注射)I.M.肌内[注射]I.U.免疫单位;国际单位I.V.静脉注射IA 免疫测定IAC 免疫亲和层析IAE 免疫亲和提取法IAHA 免疫粘附血球凝集IB 免疫印染法IC 免疫复合物ICI 国际免疫学大会ICNB 国际细菌命名委员会ICRC 国际红十字会ICT 皮内试验Icx 免疫复合物IC 脑内;皮内ID 传染病;免疫扩散;免疫缺陷病;皮内ID50 平均感染量;半数感染量IEC 离子交换层析法IEM 免疫电镜技术IEMA 免疫酶测定法IEM 免疫电镜IEOP 免疫电渗电泳IEP 免疫电泳IER 离子交换树脂IES 免疫酶染色法IF 免疫荧光IFA 免疫荧光测定IFN 干扰素IFT 免疫荧光技术Ig 免疫球蛋白IHN 抑制剂IHR 心脏固有心率II 黄疸指数IHS 人免疫血清IIF 间接免疫荧光IIP 间接免疫酶过氧化物酶(技术)IL 白细胞介素IMC 免疫记忆细胞ImD 免疫剂量ImD50 半数免疫剂量immun 免疫(接种)IMPACT 微粒计数免疫分析(法)INA 灭活剂IncB 包涵体inj 注射inoc 接种IOD 总光密度IP 免疫沉淀反应IQ 智商IR 免疫受体Ir 免疫应答IRA 免疫放射测定法IRF 免疫反应性部门IRG 免疫应答基因IRMA 免疫放射分析法iRNA 免疫核糖核酸IRU 国际参考单位IS 免疫血清ISG 免疫血清球蛋白ISH 原位杂交IS 免疫血清ISO 国际标准化组织IT 免疫毒素;皮内试验ITP 特发性血小板减少性紫癜ITT 免疫解毒疗法IU 免疫单位;国际单位IUIS 国际免疫学会联合会IUMS 国际微生物学会联合会IV 流感病毒IVI 重要值指数IVP 体外致敏的J 返回顶部JE 日本脑炎JEV 日本脑炎病毒K 返回顶部K cell杀伤细胞K 常数KA 结合常数Kaff 亲和常数Kan 卡那霉素Kc 千周KD 川崎病Kd 离解常数Keq 平衡常数Ks 底物常数L 返回顶部L.D.致死量L.F.D.最低致死量(毒素)L.R. 实验室试剂L+ 致死界量L0 无毒界量LA 乳酸;白细胞凝集素Lab 实验室LAIR 乳胶凝集抑制试验LAIT 白细胞黏附抑制试验LAI 白细胞粘附抑制试验LAK cell 淋巴因子激活的杀伤细胞LAK-CM LAK细胞条件培养基LAP 淋巴细胞激活产物LAR 迟发哮喘应答lb 磅LB 枸杞(中药)L-BCG 活卡介苗LC 致死浓度;液相层析法;活细胞LC50 半数致死浓度LCF 白细胞趋化因子LCM 淋巴细胞条件培养基LC-MS 液相层析-质谱法(联用)LCR 连接酶链反应LCT 淋巴细胞毒性试验LD 致死量LD50 半数致死量LDB 军团菌病细菌LDCC (外源)凝集素依赖性细胞介导的(细)胞毒作用LDD 最小可测出剂量法LDL 低密度脂蛋白LDMC (外源)凝集素依赖性巨噬细胞介导的(细)胞毒作用LDR 连接检测反应LDS 侧向扩散系统LE 红斑狼疮LED 最小有效剂量LEIA 发光酶免疫测定(法)LEMIT 发光酶放大免疫测定技术LEx 肝提取物Lf 絮凝单位;絮凝限度Lf 絮状反应限量;絮状反应单位LFD 最小致死量(毒素)LFT 乳胶结合试验LFU 对数荧光单位LGL 大颗粒淋巴细胞LH 乳白蛋白水解物;利度卡因;黄体生成素LI 标记指数LIA 发光免疫测定法LIBA 线形免疫结合分析法LK 淋巴因子LLC 液-液色谱法;液-液层析法LLE 液-液萃取法LM 光学显微镜术LMC 淋巴细胞介导的细胞毒性LMIT 白细胞移动抑制试验LMI 白细胞被动抑制试验LMW 低分子量LMW-UK 低分子量尿激酶LN 液氮;批号;淋巴结LNF 液氮冷冻LOD 检测极限LOH 杂合性丢失Lp 噬肺军团菌LP 停滞期;潜伏期LP 脂蛋白LpA 鲎凝集素LPA 乳胶凝集LPC 限制性前体细胞LPF 促白细胞增多因子LPL 脂蛋白脂酶LPS 脂多糖lq 液体LR 低应答者Lr 反应限量LRT 下呼吸道LS 生命科学;文献检索LSC 液体闪烁记数LSE 低盐提取物LSF 促淋巴细胞增多因子LSZ 溶菌酶LT 低温;淋巴毒素LTM 长时记忆LTS 长时间存活LTT 淋巴细胞转化试验LU 实验室单位;溶解单位LVS 活疫苗株L Y 淋巴细胞L YS 溶菌酶M 返回顶部M.E.D.最小有效量M.F.D.最小致死量M.I.D.最小感染量m.p. 熔点M.R.D.最小反应量MAb 单克隆抗体MAC (补体)膜攻击复合物MAD 平均累积剂量MAI 最大关节炎指数MAR 混合细胞凝集反应max 最大值MBR 平均结合等级MC 肥大细胞MCA 单克隆抗体McAb 单克隆抗体MCD 平均连续差MCF 巨噬细胞趋化因子MCV 平均细胞容积MDD 平均每日剂量MDO 开始反应的平均天数MED 最小有效量mel 黑色素瘤MFD 最小致死量MFI 平均荧光强度MG 重症肌无力MGC 多核巨细胞MGF 巨噬细胞生长因子mHC 次要组织相容性复合体MHC 主要组织相容性复合体MHD 维持性血液透析;最小血凝量;最小溶血量MHS 主要组织相容性系统MI 成熟指数MIA 平均积分吸光度MIC 最小抑制浓度mic 显微镜检查法Microbiol 微生物学MIET 微量免疫酶试验MIF 平均荧光强度;移动抑制因子MIg 单克隆免疫球蛋白MIHA 次要组织相容性抗原MIT 代谢抑制试验;移动抑制试验MK 猴肾(细胞培养)MLC 最小致死量;混合淋巴细胞培养MLD 半致死量MLE 最大似然估计MLF 平均线形荧光MLO 类支原体MM 恶性黑色素瘤;黏膜MMFT 微量膜荧光试验MMIT 巨噬细胞移动抑制试验mmol 毫摩尔MMR vaccine 麻疹-腮腺炎-风疹三联疫苗MN 脑膜炎球菌MND 最小坏死量mol.wt. 分子量mortal 死亡率MP 奶粉MPD 最大容许剂量MPn 支原体肺炎MPN 最大概率数MPNM 最大概率法MPS 分支杆菌多糖MPS 粘多糖MR 最大释放MRA 支原体排除剂MRD 最小反应量MRL 医疗档案室mRNA 信使RNAmRNA 信使核糖核酸MRV 混合呼吸道疫苗;混合呼吸道病毒MRU 最小识别单位MS 质谱法;分子筛MSE 均方误差MSP 微量分光光度计MSPRIA 微量固相放射免疫测定法MST 平均存活时间MT 微量滴定;结核分支杆菌MTD 最大耐受量MTX 氨甲喋呤MYO 肌红蛋白Mφ 巨噬细胞N 返回顶部N.P.N.非蛋白氮NA 核酸NAD 无异常发现NANBH 非甲非乙型肝炎NBS 新生牛血清NC 阴性对照NCM 硝化纤维膜NCS 正常鸡血清NFD 非致死性疾病NHP 正常人血浆NHG 正常人球蛋白NHS 标准人血清NI 中和指数NIM 神经免疫调节NK cell 正常杀伤细胞NKDA 不明药物过敏NLE 正常肝浸出物Nm 脑膜炎奈瑟氏球菌NMR 核磁共振NMS 正常小鼠血清NOAEL 无明显副作用剂量NPC 鼻咽癌NPN 非蛋白氮nRNA 正常核糖核酸NPO 禁食NRP 正常兔血清NRS 正常家兔血清NS 生理盐水;神经系统;正常血清NSS 非免疫绵羊血清NS 生理盐水;正常血清NT 中和试验O 返回顶部OD光密度ODU 光密度单位OI 调理指数OM 外膜Op 手术O-PS 菌体多糖OPV 口服脊髓灰质炎病毒疫苗OS 低聚糖;寡糖OUR 摄氧率Ova 卵巢OT 旧结核菌素P 返回顶部P.H.公共卫生p.i. 腹腔注射;接种后PA 恶性贫血pAB 多克隆抗体PAGE 聚丙烯酰胺凝胶PAGE 聚丙烯酰胺凝胶电泳pat. 专利PB 外周血PBS 磷酸盐缓冲盐水PC 吞噬细胞;PCE 纸层析电泳法PCP 肺炎球菌荚膜多糖PCR 蛋白分解率Pep 胃蛋白酶petro. 液体石蜡pf 无热原的PFU 空斑形成单位PFU 空斑形成单位;疱斑形成单位PHA 植物血凝素PHAI 被动血球凝集抑制PHA 被动血球凝集;植物血凝素PHC 原发性肝癌Phys 生理盐水pI 等电点PI 增殖指数PMS 孕马血清ppt 千分之一;沉淀物prev 预防PS 病人血清PSE 猪脾提取物PSE 丙酮醛固定的绵羊红细胞PSS 生理盐溶液PT 百日咳毒素PVC 聚氯乙烯PW 石蜡Py 嘧啶Pyr 吡啶Q 返回顶部q.v. 参照QA 质量保证QC 质量控制Q-F 速冻法QM 质量管理QO2 氧商QS 质量体系QT 快速试验R 返回顶部R 受体R.B.C.红细胞(旧名红血球)RAID 放射免疫检测法RAM 微生物放射测定法RBC 红细胞计数RC 红十字会RCA 红细胞凝集RD 风湿病Ref 参考文献RFP 利福平RFT 花环形成试验RFFIT 狂犬病荧光聚焦抑制试验RH 相对湿度rh 重组人的rhEPO 重组人红细胞生成素rhG-CSF 重组人粒细胞集落刺激因子rhIL-2 重组人白细胞介素-2RIAGDA 放射免疫琼脂凝胶扩散自显影RIAT 放射免疫吸收试验RIA 放射免疫自显影;放射免疫分析RICEPA 放射免疫对流电泳自显影RICR 红细胞免疫复合物花结形成率RIE 火箭免疫电泳RIEP 火箭免疫电泳RIST 放射免疫吸附试验RIT 花环抑制试验RM 标准物质RNA 核糖核酸RNase 核糖核酸酶RNA 核糖核酸RPHA 反向被动血凝rpm 每分钟转数RQ 呼吸商rRNA 核糖体核糖核酸RRP 相对不应期RSA 兔血清白蛋白RV 轮状病毒S 返回顶部Sa 金黄色葡萄球菌S.A.S. 统计分析系统SARS 重度急性呼吸综合征,非典型肺炎s.v.酒精;乙醇SCAT 羊红细胞凝集试验SCL 硬皮病SD 标准差SEA 绵羊红细胞凝集反应SEM 均值标准误差SEV 副流感病毒SE 标准误差(统计)SFDA State DrugAdiministration 国家食品药品监督管理局SFI 特异性荧光强度SH 重症肝炎SH.血清性肝炎SM 合成培养基Sm 链霉素Sn 上清液SNS 交感神经系统SOD 超氧化物歧化酶SIT 血清抑制试验SP 单阳性sp.act. 比活度SPE 固相提取SPT 皮刺试验SPV 合成肽疫苗SR 沉降率SRBC 绵羊红细胞SRID 单项放射免疫扩散sRNA 可溶性核糖核酸SS 唾液标本SSA 表面结构分析SSPE 亚急性性硬化性全脑炎ST 皮肤试验ster 灭菌,绝育STD 性传播疾病;标准试验剂量strep 链球菌;链球菌属STH 生长激素STT 致敏试验STU 皮肤试验单位SV40 猴病毒四十号;空泡病毒SYN 合成率T 返回顶部T 靶细胞T.A.T. 毒素抗毒素;破伤风抗毒素T.b.结核杆菌t1/2 半衰期TAA 总氨基酸;肿瘤相关抗原TABC vaccine 伤寒-副伤寒甲、乙、丙菌苗TAT 破伤风抗毒素TAT 吐温-白蛋白-磷酸缓冲液Tbc 结核杆菌TBM 结核性脑膜炎TB 结核菌素Tc 细胞毒性T细胞TC 组织培养TCA 三氯乙酸TCD50 半数组织培养量TCF 双色荧光法TCGF T 细胞生长因子TCID50 50%组织培养感染量TCID50 半数组织培养感染量TD50 半数中毒量Te 破伤风Tet 破伤风类毒素TF 冷冻温度;转移因子TFU 转化单位Tk 杀伤T细胞TID50 半数组织感染量TIF 肿瘤抑制因子TLT 结核菌素划痕试验TM 胸腺素TNF 肿瘤坏死因子TO 靶器官TP 胸腺生成素TR 总释放;转化率;结核菌素反应tRNA 转移核糖核酸TS 三染法TSA 肿瘤特异性抗原TT 破伤风类毒素TU 毒素单位TWCC 总白细胞计数U 返回顶部U.S.Pat. 美国专利UF 超滤法UHT 超高温ULD 超低剂量UK 尿激酶Ura 尿嘧啶US 超声波检查法UVF 紫外荧光UVR 紫外线V 返回顶部V 疫苗V.S.滴定[用] 溶液;定量溶液Var 方差VC 水痘VCG 卡介苗Ve 洗脱体积VG 黏度梯度VHF 病毒性出血热Vi 毒力VMC 病毒性心肌炎VNT 病毒中和试验VV 牛痘病毒vv 静脉VZV 水痘带状疱疹病毒W 返回顶部W.B.C. 白细胞;白细胞计数WB 全血WCV 全细胞疫苗WHO 世界卫生组织WHS 人全血清WM 洗涤剂WSC 全脾细胞X-Z 返回顶部X matching 交叉配血XR X射线YC Y染色体YFV 黄热病毒ZE 区带电泳ZIG 带状疱疹免疫球蛋白其它返回顶部αFP 甲胎蛋白α-Gal 半乳糖γ-globulin 丙种球蛋白。
adpr分子式

adpr分子式ADPR(Adenosine diphosphate ribose)是一种重要的分子式,它由腺苷二磷酸(ADP)和核糖(ribose)组成。
ADPR分子式的重要性在于它在生物体内发挥着多种重要的生理功能。
ADPR分子式在细胞能量代谢中起着重要作用。
ADP是一种能量的储存形式,当细胞需要能量时,ADP会通过磷酸化反应转化为三磷酸腺苷(ATP),释放出能量供细胞使用。
而ADPR则是ADP在细胞内的中间产物,它在细胞内能量代谢过程中起到了催化反应的作用。
通过ADPR的参与,细胞能够高效地将ADP转化为ATP,从而维持细胞正常的能量供应。
除了在能量代谢中的作用外,ADPR分子式还与细胞信号转导有关。
在细胞内,ADPR可以通过与其他蛋白质结合来调节细胞内信号传递的过程。
例如,ADPR可以与钙离子结合,影响钙离子在细胞内的浓度,从而调节细胞的生理功能。
此外,ADPR还可以与一些酶结合,调控细胞内的酶活性,进而影响细胞的代谢过程和生物学功能。
ADPR分子式还在细胞增殖和凋亡过程中发挥着重要作用。
ADPR可以与细胞内的一些蛋白质结合,调节细胞的增殖和凋亡信号通路。
当细胞需要增殖时,ADPR可以通过激活细胞周期蛋白依赖激酶(CDK)促进细胞周期的进行。
而当细胞需要凋亡时,ADPR可以激活凋亡信号通路中的关键酶,引导细胞进入凋亡过程。
ADPR分子式还与DNA修复过程有关。
当DNA受到损伤时,细胞会通过一系列复杂的修复机制来修复DNA。
而ADPR则可以与DNA修复酶结合,促进DNA修复过程的进行。
通过与DNA修复酶的结合,ADPR 可以增强酶的活性,加快DNA修复的速度,从而维护细胞的基因稳定性。
ADPR分子式在细胞内发挥着多种重要的生理功能。
它在细胞能量代谢、细胞信号转导、细胞增殖和凋亡以及DNA修复等过程中起着重要作用。
对于了解细胞生物学和疾病发生机制,研究ADPR的生物学功能具有重要意义。
希望在未来的研究中,我们能够更深入地探索ADPR的功能和机制,为人类健康和疾病治疗提供更多的突破。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
货号:MS3403 规格:100管/96样腺苷二磷酸葡萄糖焦磷酸化酶(ADP-glucose pyrophosphorylase,AGP)
活性测定试剂盒说明书
微量法
正式测定前务必取2-3个预期差异较大的样本做预测定
测定意义:
AGP(EC 2.7.7.21)主要存在于植物中,催化葡萄糖-1-磷酸与ATP反应生成淀粉合成的直接前体ADPG,是植物淀粉生物合成的主要限速步骤。
测定原理:
AGP催化的逆向反应生成G1P,在反应体系中添加的磷酸己糖变位酶和6-磷酸葡萄糖脱氢酶依次催化生成6-磷酸葡萄糖酸和NADPH,340nm下测定NADPH增加速率,即可计算AGP活性。
自备实验用品及仪器:
紫外分光光度计/酶标仪、水浴锅、台式离心机、可调式移液器、微量石英比色皿/96孔板、研钵、冰和蒸馏水
试剂的组成和配制:
提取液:液体100mL×1瓶,4℃保存;
试剂一:液体20mL×1瓶,4℃保存;
试剂二:粉剂×1瓶,4℃保存;临用前加入6.4mL蒸馏水充分溶解备用;用不完的试剂分装后-20℃保存,禁止反复冻融;
试剂三:粉剂×1瓶,-20℃保存;临用前加入4.8mL蒸馏水充分溶解备用;用不完的试剂分装后-20℃保存,禁止反复冻融;
粗酶液制备:
按照组织质量(g):提取液体积(mL)为1:5~10的比例(建议称取约0.1g组织,加入1mL 提取液),进行冰浴匀浆。
10000g 4℃离心10min,取上清,置冰上待测。
测定步骤:
1、分光光度计或酶标仪预热30min以上,调节波长至340nm,蒸馏水调零。
2、试剂一置30℃保温10min以上。
3、在EP管中按顺序加入下列试剂(如果一次性测定样本较多,可以将试剂一和试剂二按比例
2)
A1
第1页,共2页
和 2min后的吸光度A2,计算ΔA=A2-A1。
AGP活性计算:
a.使用微量石英比色皿测定的计算公式如下:
1、按样本蛋白蛋白浓度计算
单位的定义:每mg组织蛋白每分钟催化产生1nmol NADPH定义为一个酶活性单位。
AGP(nmol/min/mg prot)=[ΔA×V反总÷(ε×d)×109]÷(V样×Cpr) ÷T
=2813×ΔA÷Cpr
此法需要自行测定样本蛋白质浓度。
2、按照样本鲜重计算
单位的定义:每g组织每分钟催化产生1nmol NADPH定义为一个酶活力单位。
AGP(nmol/min /g 鲜重)=[ΔA×V反总÷(ε×d)×109]÷(W× V样÷V样总)÷T
=2813×ΔA÷W
V反总:反应体系总体积,2.8×10-4 L;ε:NADPH摩尔消光系数,6.22×103 L / mol /cm;d:比色皿光径,1cm;V样:加入样本体积,0.008 mL;V样总:加入提取液体积,1 mL;T:反应时间,2 min;Cpr:样本蛋白质浓度,mg/mL;W:样本质量。
b.使用96孔板测定的计算公式如下:
1、按样本蛋白蛋白浓度计算
单位的定义:每mg组织蛋白每分钟催化产生1nmol NADPH定义为一个酶活性单位。
AGP(nmol/min /mg prot)=[ΔA×V反总÷(ε×d)×109]÷(V样×Cpr) ÷T
=5626×ΔA÷Cpr
此法需要自行测定样本蛋白质浓度。
2、按照样本鲜重计算
单位的定义:每g组织每分钟催化产生1nmol NADPH定义为一个酶活力单位。
AGP(nmol/min /g 鲜重)=[ΔA×V反总÷(ε×d)×109]÷(W ×V样÷V样总)÷T
=5626×ΔA÷W
V反总:反应体系总体积,2.8×10-4 L;ε:NADPH摩尔消光系数,6.22×103 L / mol /cm;d:96孔板光径,0.5cm;V样:加入样本体积,0.008 mL;V样总:加入提取液体积,1 mL;T:反应时间,2 min;Cpr:样本蛋白质浓度,mg/mL;W:样本质量。
第2页,共2页。