第2章基因工程的酶学基础
2基因工程的酶学基础

GCGC NarI BsaHI
GGCC
BbeI HaeII XbaI Bcl I
FspI
ApaI BanII
Bsp1286
T****A T****A NruI T****A T****A
EaeI MscI
GTAC **** **** **** **** A****T A****T A****T A****T A****T
② 不完全同裂酶: 识别位点相同,但切点不同。 如Xma I 和 Sma I。
Xma I
Sma I
5’-CCCGGG -3’ 3’-GGGCCC-5’ 5’-CCCGGG-3’ 3’-GGGCCC-5’
(6)同尾酶(Isocaudamers)
识别的序列不同,但能切出相同的粘性末 端。如BamH I、Bgl Ⅱ、Bcl I、Xho Ⅱ等
核酸限制性内切酶的类型及主要特性
特性 I类内切酶 II类内切酶 III类内切酶
限制和修饰 单一多功能的酶 活性 内切酶和甲基 共同亚基的双 化酶分开 功能酶
内切酶的蛋 3种不同的亚基 白结构 限制作用所 ATP、 Mg2+和SAM 需要的辅助 因子 寄主特异性 EcoB: 位点识别序 TGA(N)8TGCT 列 EcoK: AAC(N)6GTGC
BamH I Bgl Ⅱ
Bcl I Xho Ⅱ
5’-GGATCC-3’ 3’-CCTAGG-5’
5’-AGATCT-3’ 3’-TCTAGA-5’ 5’-TGATCA-3’ 3’-ACTAGT-5’ 5’-UGATCY-3’ 3’-YCTAGU-5’
U代表嘌呤;Y代表嘧啶。
Sau 3A
BfaI
DpnI
HinpI
HaeIII HhaI
第二章兰州大学基因工程基因工程酶学基础

(2)切割位点
•绝大多数II类酶在其识别位点内切割 DNA
PPT文档演模板
第二章兰州大学基因工程基因工程酶 学基础
• 粘性末端 (cohensive end)
• 连接便 利
PPT文档演模板
第二章兰州大学基因工程基因工程酶 学基础
PPT文档演模板
第二章兰州大学基因工程基因工程酶 学基础
• DNA 分子末端标 记
PPT文档演模板
•Dr. Werner Arber
第二章兰州大学基因工程基因工程酶 学基础
•2 性 质
内切酶,即在核酸分子内部制造切 口的酶
形成5`-P 和3`-OH末端
•3 功 能
• 自我保护作用 • 细菌的限制和修饰系统(R/M体系)
PPT文档演模板
第二章兰州大学基因工程基因工程酶 学基础
a 限制(Restriction)
• 大多数的核酸限制内切酶的标准反应温度 为37 ℃ ,但也有许多例外,如SmaI为25 ℃ ,ApaI为30 ℃ ,TaqI为65 ℃
c 限制修饰系统分子机理
• 由三个连续基因位点所控制:
hsd R, hsdM, hsd S
• hsd R---限制性内切酶:能识别DNA分子 上的特定位点并将双链DNA切断
• hsd M---限制性甲基化酶 :催化DNA分子 特定位点上的碱基甲基化反应
• hsd S---控制两个系统的表达 :协助上述 两种酶识别特殊的作用位点
• 1、DNA的纯度 • 2、DNA的甲基化程度 • 3、酶切反应的温度 • 4、DNA的分子结构 • 5、核酸限制性内切酶的反应缓冲液
PPT文档演模板
第二章兰州大学基因工程基因工程酶 学基础
DNA的纯度
基因工程-酶工程-细胞工程重点

目录基因工程重点 (1)第1章绪论 (1)第2章基因工程的酶学基础 (1)第3章基因工程载体 (3)第4章基因工程的主要技术及其原理 (4)第5章目的基因的获得 (7)第6章DNA重组的操作 (8)第7章外源基因的表达及其优化策略 (8)酶工程重点 (13)第一章绪论 (13)第二章酶工程基础 (13)第三章酶的发酵工程 (15)第四章酶的分离工程 (16)第五章固定化酶和固定化细胞 (18)第六章化学酶工程 (19)第七章非水相酶催化 (20)第八章核酶 (22)第十章酶抑制剂 (22)第十一章酶的应用 (23)细胞工程重点 (25)第一章绪论 (25)第四章细胞培养 (25)第五章细胞融合与单克隆抗体 (27)第六章胚胎工程 (28)第七章干细胞与组织工程 (29)第八章核移植技术和动物克隆 (31)第九章转基因动物和生物反应器 (32)第十章动物染色体工程 (34)第十一章植物组织培养 (34)第十二章植物的快速繁殖 (36)I第十三章单倍体诱导和育种 (37)第十四章植物胚胎培养 (38)第十五章体细胞胚发育和人工种子 (38)第十六章植物原生质体融合技术 (39)第十七章植物染色体工程 (41)第十八章植物转基因技术 (42)II基因工程重点第1章绪论一、基因工程概念(Conception ):基因工程(gene engineering):就是对不同生物的遗传物质,在体外进行剪切、组合和拼接,使遗传物质重新组合,然后通过载体转入微生物、植物和动物细胞内,进行无性繁殖,并使所需要的基因在细胞中表达,产生出人类所需要的产物或组建成新的生物类型。
二、基因工程一般操作步骤:1、目的既赢得获取2、基因表达载体的构建3、将目的基因导入受体细胞4、目的基因的检测和表达产物的测定三、基因工程四大要素(Four elements):1、供体基因2、载体3、工具酶4、受体细胞(细菌、植物和动物)第2章基因工程的酶学基础一、名词解释:1.限制/修饰系统(R-M, Restriction-modification system)——任何一种生物体都存在防御外界物质进入的机制。
工学基因工程第2章酶学基础2DNA连接酶

• 黏性末端修饰成平末端后再连接
3’突出末端
DNA片段末端同聚物加尾后连接
• 所谓连杆(1inker),是指用化学方法合成的一 段由10~12个核苷酸组成的、具有一个或数个 限制酶识别位点的寡核苷酸片段。
• DNA片段末端加上化学合成的连杆,能形成黏 性末端,连接后可用内切酶进行切割。
• 连杆的5’-末端和待克隆的DNA片段5’末端,用多核苷酸激酶处理使之磷酸 化,然后再通过T4DNA连接酶的作用 使两者连接起来。接着用适当的限制 酶消化具有连杆的DNA分子和克隆载 体分子,这样的结果使二者都产生出 了彼此互补的黏性末端。于是便可以 按照常规的黏性末端连接法,将待克 隆的DNA片段同载体分子连接起来。
nick P OH
DNA连接酶
5‘…G--C--T--C--T--G--C--A--G--G--A--G … 3’
3‘…C--G--A--G--A--C--G--T--C--C--T--C … 5’
• 修复与RNA链结合的DNA链上切口处的磷 酸二酯键
5‘…G--C--U--C--U--G--C--A--G--G--A--G … 3’
这种方法的优点是定向连接克隆,筛选方便。 缺点则是PCR产物的酶切和判断比较困难,另 外酶切连接过程本身也相当的费时、费力。
现象:普通的Taq酶会自动给PCR产物的3’末端添 加一个A(3’ -A) ,因此不能直接将PCR产物末 端抹平后连接。
解决方法: • Fill in the ends with Klenow(Klenow酶处理PCR产物将
基因工程的酶学基础

DNA甲基化对酶活性的影响 •大多数大肠杆菌菌株中含有Dam甲基化酶 和Dcm甲基化酶。 •Dam可以在GATC序列中腺嘌呤N-6位上 引入甲基 •Dcm在CCA/TGC序列的第一个胞嘧啶C-5 位置上引入甲基。 •部分限制性内切酶对甲基化的DNA不能切 割,如FbaI和MboI等。
通常有两种方法: ①选用上述酶的同功酶,如Sau3AI,DNA 识别切割位点与MboI相同;但不受甲基化 影响; ②利用甲基化酶缺失的受体细胞进行DNA 的制备,如E. coli & nbsp; JM110和链霉菌 等,前者Dam和Dcm甲基化酶已敲除,而 后者细胞内本就没有甲基化酶,从这些细 胞中抽提的DNA就能被上述酶切割。
CATG
SAGE Tag 5
CATG
SAGE Tag 2
SAGE Tag 4
SAGE Tag 6
•II型限制性内切酶的酶切位点位于识别位点之中或
附近,产生以下4种情况。 ① 5’粘性末端;② 3’粘性末端;③ 平末端和 ④ 非互补的粘性末端 •无论DNA的来源如何,只要是使用同一种限制性内 切酶,所留下的残端都是一样的。 •经酶切后,5’端总是保留一个磷酸根;3’端总是 保留一个OH
第二讲 基因工程的酶学基础
本节内容
•限制性内切酶—主要用于DNA分子的特异切割 •DNA甲基化酶—用于DNA分子的甲基化 •核酸酶—用于DNA和RNA的非特异性切割 •核酸聚合酶—用于DNA和RNA的合成 •核酸连接酶—用于DNA和RNA的连接 •核酸末端修饰酶—用于DNA和RNA的末端修饰 •其它酶类--用于生物细胞的破壁、转化、核酸纯化、 检测等。
Ⅱ型限制性内切核酸酶的基本特性
1.具有高度特异性的识别位点与酶切位点。 2.辅基: Mg++。 特定识别序列一般长4 - 8对碱基; 大部分酶的切割位点在识别序列内部或两侧;
[医学保健]基因工程的酶学基础
![[医学保健]基因工程的酶学基础](https://img.taocdn.com/s3/m/01de5ec902768e9950e73888.png)
碱性磷酸酶:
来自于大肠杆菌(bacterial alkaline phosphatase BAP)或小牛肠(calf intestinal alkaline phosphatase CIP),该酶用于脱去 DNA(RNA)5’末端的磷酸根,使5’-P成为5’-OH,该 过程称核酸分子的脱磷酸作用。当需要5’端同位 素标记或为了避免DNA片段自身连接(或环化)时 可进行脱磷酸反应。
该酶全称为末端脱氧核苷酸转移酶(terminal deoxynucleotidyl transferase),它是从小牛胸腺中 纯化出来的一种小分子量的碱性蛋白质,在二甲胂 酸缓冲液中,能催化5’脱氧核苷三磷酸进行5’-3’ 方向的聚合作用,逐个地将脱氧核苷酸分子加到线 性DNA分子的3’-OH末端。
精品课件文档,欢迎下载。下 载后可以复制、编辑。
上个世纪末,本世纪初,1996年,清华学界 对中医 气本质 ,经络 实质, 阴阳, 五行, 藏象, 中医哲 学观等 都有了 新的全 面整体 创造性 的认识 和解说 。如, 邓宇等 发现的 :气是 流动着 的‘信 息-能 量-物 质’的 混合统 一体; 分形分 维的经 络解剖 结构; 数理阴 阳;中 医分形 集:分 形阴阳 集-阴 阳集的 分形分 维数, 五行分 形集- 五行集 的分维 数;分 形藏象 五系统 -暨心 系统、 肝系统 、脾系 统、肺 系统、 肾系统 ;中医 三个哲 学观- 新提出 的第三 哲学观 :相似 观-分 形论等 。
3’CTTAAG 5’
平末端
两条链上的断裂位置是处在一个 对称结构的中心这样形式的断裂是 形成具有平末端的DNA片断。不易 重新环化。
同裂酶(isoschizomers)
指来源不同但识别相同靶序列的核酸内 切酶。同裂酶进行同样的切割,产生同样的 末端。但有些同裂酶对甲基化位点的敏感性 不同。 Example: 限制酶HpaⅡ和MspⅠ是一对同裂酶 (CCGG) , 当靶序列中一个5-甲基胞嘧啶时HpaⅡ不能进行 切割,而MspⅠ可以。
基因工程的酶学基础PPT课件

本名称。
大肠杆菌(Escherichia coli)用Eco表示; 流感嗜血菌(Haemophilus influenzae)用Hin表示
6
第6页/共105页
4. 如果酶存在于一种特殊的菌株中,则将该菌株 名的第一个字母加在基本名称后,若酶的编码基因 位于噬菌体(病毒)或质粒上,则用一个大写字母 表示此染色体外遗传成分。
ii)同一个DNA分子内连接: 通过两个相同的粘性末端可以连接成环 形分子。
21
第21页/共105页
② 末端标记
A 末端可用DNA多核苷酸激酶进行32P标记。 B 末端可以通过末端转移酶添加几个多聚核苷酸 的尾巴(如dA和dT),即同聚物加尾造成人工粘 性末端。
③ 补平成平齐末端
粘性末端可以用DNA聚合酶补平成平齐末端。
22
第22页/共105页
[3] 平齐末端(blunt end)
在识别序列的对称轴上同时切割 如EcoR V
5’-
GATATC
-3’
3’-
CTATAG
-5’
23
第23页/共105页
[4] 平齐末端的特点
连接困难,连接效率低,只有粘性末端连接 效率的1%,这种连接常出现多联体连接。
粘性末端与平齐末端连接的处理方法
-3’ -5’
19
ⅱ) 3’端凸出(如Pst I切点)
5’-
CTGCAG
-3’
3’-
GACGTC
-5’
5’-
CTGCA
G
-3’
3’-
G
ACGTC
-5’
基因工程 第二章 基因工程的酶学基础

GATC CTAG SauI G TCGAC CAGCTG
BamH I
GGATCC CCTAGG Xho I CTCGAG GAGCT C GTCGAG
?
CAGCTC
利用同尾酶可使切割位点的选择余地更大(相容性末端);
同时,同尾酶产生的DNA片段能通过粘性末端之间的互 补作用而彼此连接起来,这在分子克隆中有一定的作用;
(三)DNA的甲基化程度 原核生物细胞内存在限制-修饰系统,其中MTase 甲基转移酶对DNA起修饰作用,从而使自身DNA不 受体内限制性内切核酸酶的切割 甲基化作用直接影响限制性内切核酸酶的活性 为了克服MTase甲基转移酶的影响,在基因克隆 中使用MTase缺失的菌株来制备质粒DNA。
普遍采用的是Smith和Nathans(1973)提议的 命名系统 由属名的第一个字母(大写)和种名的前两个字 母(小写),组成3个字母的略语组成酶的基本 名称。
例如,大肠杆菌(Escherichia coli)用Eco表示, 流感嗜血菌(Haemophilus influenzae)用Hin表示。
2 3 酶的纯度 DNA样品的纯度 DNA的甲基化程度
5
6 3 7
DNA分子的构型 反应缓冲液 酶的星号活性
4
酶切反应的温度
(一)酶的纯度 高质量的限制性内切核酸酶应是通过全面 提纯的,不存在其他内切核酸酶或外切酶的 污染
在长时间酶解时不出现识别序列特异性的下降 酶解的DNA片段连接后能重新被识别和切割等。
Ry13
EcoR I
属名 种名 株系 编号
• 若种名头2个字母相同则其中一个可用种名的第一和第三个 字母。
Escherichia coli RI — EcoRI Haemophilus influensae d Ⅲ— HindⅢ Streptomyces achromagenes I (Ⅱ) — SacI (II)
基因工程的酶学基础

切割方式
Ⅱ限制性核酸内切酶切割双链DNA,水解磷酸二酯键中3’ 位酯键产生两个末端,末端结构是5’-P和3’-OH,产生3种不同的切口。
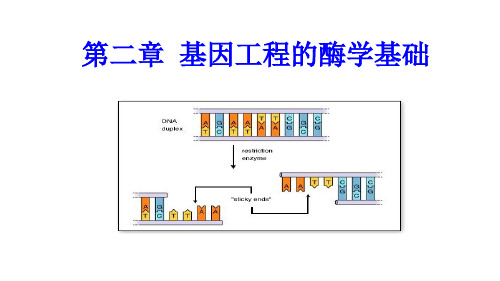
粘性末端(cohesive ends):因酶切位点在两条DNA单链上不同(对称)酶切后形成的具有互补碱基的单链末端结构,酶切产生的两个粘性末端很容易通过互补碱基的配对而重新连接起来。
第二章 基因工程的酶学基础
从核酸分子末端逐个降解核苷酸,叫外切核酸酶 从核酸分子内部切割磷酸二酯键使之断裂形成小片段,叫内切核酸酶
按水解断裂核酸分子的方式:
核酸酶:通过切割相邻的两个核苷酸残基之间的磷酸二酯键,从而导致核酸分子多核酸链发生水解断裂的蛋白酶。 特异水解断裂RNA分子,核糖核酸酶(RNase) 特异水解断裂DNA分子,脱氧核糖核酸酶(DNase) 非专一性酶,底物或者为DNA或者为RNA
主要特性
Ⅰ型
Ⅱ型
Ⅲ型
限制修饰 蛋白结构 辅助因子 识别序列 切割位点
双功能(具甲基化) 异源三聚体 ATP Mg2+ SAM 距识别序列1kb处 随机性切割
单一功能 同源二聚体 Mg2+ 4-6bp回文序列 识别序列内或附近特异切割
双功能(具甲基化) 异源二聚体 ATP Mg2+ 距识别序列下游 24-26bp处 随机性切割
01
大肠杆菌(Escherichia coli)用Eco表示; 流感嗜血菌(Haemophilus influenzae)用Hin表示
01
限制性内切酶的命名
如果酶存在于一种特殊的菌株中,则将该菌株名的第一个字母加在基本名称后,若酶的编码基因位于噬菌体(病毒)或质粒上,则用一个大写字母表示此染色体外遗传成分。
第二章基因工程的酶学基础精品文档29页

幻灯片1第二章基因工程的酶学基础Enzymes幻灯片2第一节限制性核酸内切酶一、限制性核酸内切酶(Restriction endonuclease)是一类能够识别双链DNA分子中的某种特定核苷酸序列(4—8bp),并由此处切割DNA双链的核酸内切酶。
幻灯片31.来源原核生物。
2. 性质内切酶。
即在核酸分子链的内部制造切口的酶。
3.功能自我保护作用。
细菌的限制和修饰系统(R/M体系)幻灯片4(1)限制(R e s t r i c t i o n)限制性内切酶将侵入细菌体内的外源DNA切成小片断。
幻灯片5(2)修饰(M o d i f i c a t i o n)细菌自身的DNA碱基被甲基化酶甲基化修饰所保护,不能被自身的限制性内切酶识别切割。
① Dam甲基化酶GATC 腺嘌呤N6位置引入甲基② Dcm甲基化酶CCAGG或CCTGG序列在第二个C上C5位置上引入甲基幻灯片6幻灯片7二、限制性内切酶的类型目前鉴定出三种不同类型的限制性内切酶。
I 型限制性内切酶I I类限制性内切酶I I I类限制性内切酶幻灯片81.I型限制性内切酶首先由M. Meselson和R. Yuan在1968年从大肠杆菌 B株和 K株分离的。
如 EcoB和 EcoK。
(1)识别位点序列未甲基化修饰的特异序列。
EcoB: TGA(N)8TGCTEcoK:AAC(N)6GTGC幻灯片9(2)切割位点在距离特异性识别位点约1000—1500 bp处随机切开一条单链。
Recognize sitecut1-1.5kb(3)作用机理需ATP、Mg2+和SAM(S-腺苷蛋氨酸)。
幻灯片102.I I类限制性内切酶首先由H.O. Smith和K.W. Wilcox在1970年从流感嗜血菌中分离出来。
分离的第一个酶是Hind Ⅱ(1)识别位点序列未甲基化修饰的双链DNA上的特殊靶序列(多数是回文序列)。
与DNA的来源无关。
幻灯片11(2)切割位点识别位点处。
第二章基因工程的酶学基础
第二章基因工程的酶学基础内容一、概述二、限制性内切核酸酶三、DNA连接酶四、其他工具酶一、概述工具酶:在生物技术中常用的各种工具酶系指能用于DNA和RNA分子的切割、连接、聚合、反转录等有关的各种酶系统称为工具酶。
工具酶名称主要功能限制性内切核酸酶在DNA分子内部的特异性的restriction endonucleases 碱基序列内部进行切割DNA连接酶将两条以上的线性DNA分子或片段DNA ligase 催化形成磷酸二酯键连接成一个整体DNA聚合酶I通过向3’端逐一增加核苷酸以填补双链DNA分子上的单链DNA polymerase I 裂口,即5’→3’DNA聚合酶活性与3’→5’及5’→3’外切酶活性多核苷酸激酶催化将把一个磷酸分子加到多核苷酸链的DNA polymerase kinease 5’-OH末端上(接下表格)工具酶名称主要功能反转录酶以RNA分子为模板合成互补的cDNA链reverse transcriptaseDNA末端转移酶将同聚物尾巴加到线性双链或单链DNA分子的3’-OH DNA terminal transferase 末端或DNA的3’-末端标记dNTP碱性磷酸酶去除DNA,RNA,dNTP的5’磷酸基团BAP orCIAP核酸外切酶III 降解DNA3’-OH末端的核苷酸残基exonuclease III降解酶S1 降解单链DNA或RNA,产生带5’磷酸的单核苷酸或nuclease S1寡聚核苷酸,同时也可切割双链核酸分子的单链工具酶名称主要功能核酸酶Bal 31 降解双链DNA,RNA的5’及3’末端,nuclease Bal31Taq DNA聚合酶能在高温(72℃)下的单链DNA为模板,Taq DNA polymerase从5’→3’方向合成新生的互补链核糖核酸酶专一性降解RNARNase脱氧核糖核酸酶内切核酸酶,水解单链或双链DNA DNase二、限制性内切核酸酶1、限制性核酸内切酶的分类2、限制性核酸内切酶的命名原则3、限制性核酸内切酶的基本特征4、影响限制性内切酶的活性因素5、限制性核酸内切酶的应用1、限制性核酸内切酶的分类限制性核酸内切酶主要分成三大类:I类:能识别专一的核苷酸顺序,并在识别点附近的一些核苷酸上切割双链,但切割序列没有专一性,是随机的。
基因工程的酶学基础

基因工程的酶学基础
核酸限制性内切酶的类型及主要特性
特性
Ⅰ类内切酶
Ⅱ类内切酶 Ⅲ类内切酶
限制和修 饰活性
内切酶和甲基 共同亚基的双功
单一多功能的酶 化酶分开
能酶
内切酶的 蛋白结构
3种不同的亚基
2种不同的亚基 单一成分
限制作用 所需要的 辅助因子
寄主特异 性位点识 别序列
ATP、 Mg2+和SAM
EcoB:TGA(N)8TGCT EcoK:AAC(N)6GTGC
• 2. 性
•内切酶
•即质在核酸分子链的内部制造切口的酶。
•形成5’-P和3’-OH末端
• 3. 功
•自我保护作用
能
•细菌的限制和修饰系统(R/M体系)
PPT文档演模板
基因工程的酶学基础
PPT文档演模板
基因工程的酶学基础
(1)限制(Restriction)
•限制性内切酶将侵入细菌体内的外源DNA 进行分解,切成小片断。
•如 Hind Ⅱ:d菌株 • EcoR I :抗药性R质粒
•
PPT文档演模板
基因工程的酶学基础
•3. 如果一种特殊的菌株内有几种不同的限制与 修复系统,用罗马字母表示该菌株中发现某种酶
的先后次序。
•
如Hind Ⅱ:d菌株中发现的第二个酶
•4. 所有的限制酶,除以上名称外还要冠以系统名 称。限制性内切酶的系统命名为R,甲基化酶为M。
•含有几个核苷酸单链的末端。 •分两种类型:
• ⅰ) 5’端凸出(如EcoR I切点)
•5’•3’-
•GAATTC •CTTAAG
•-3’ •-5’
•5’•3’-
PPT文档演模板
第二章兰州大学基因工程基因工程酶学基础

• 二 、特点
• 2 连接条件
• 3 DNA连接酶连接反应条件
• 三、连接反应机理
• 四、连接反应温度
• 五、影响因素
• 六、反应体系
• 七 、连接操作
• 八、体外连接DNA片段的几种方法
Nathan和Dr. Hamilton Smith博士第一次从 大肠杆菌中提取出限制性内切核酸酶 • 已经分离提取出500多种限制性内切核酸酶
Dr. Werner Arber
2性质
内切酶,即在核酸分子内部制造切 口的酶
形成5`-P 和3`-OH末端
3功能
自我保护作用 细菌的限制和修饰系统(R/M体系)
阿尔伯(Arber)、史密斯(Smith)和内森斯 (Nathans),获1978年诺贝尔生理学和医学奖
• 同裂酶 (isoschizomers): 来源不同, 识别的是同样的核苷酸靶序列
• 同尾酶(isocaudamer) :来源不同,识别的靶 序列也各不相同,但都能产生相同的粘性末端
由同尾酶所产生的DNA片段,是能够通过其粘性末端之间的 互补作用而彼此连接起来的,在基因克隆实验中很有用处
• 消化反应温度高于或低于最适反应温度, 都会影响酶的活性
• 每ugDNA 用1-5U酶切1-2h
DNA的分子结构
• 某些核酸内切酶切割超螺旋的质粒DNA或病毒DNA所需的 酶量比消化线性DNA高出20倍
• 一些核酸内切酶,切割处于不同位置的限制位点,其效率有 明显的差异, 可能是由于侧翼序列的核苷酸成分的差异造 成的
• 由一对同尾酶分别产生的粘性末端共价结 合形成的位点,称之为“杂种位点 ”(hybrid site)。但这类杂种位点的结构, 一般不再被原来的任何一种同尾酶所识别
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
限制性内切酶
DNA连接酶
DNA聚合酶和反转录酶
DNA修饰酶
外切核酸酶
单链内切核酸酶
RNA酶
核酸酶:水解相邻的两个核苷酸残基之间 的磷酸二酯键,从而使核酸分子断链。 根据水解的不同方式,分为:
外切核酸酶(exonuclease) 内切核酸酶(endonuclease)
5' …C TGCA▼G…3' 3' …G▲ACGT C…5' Pst I 5' …CTGCA3’ G…3' 3' …G 3’ACGTC… 5'
(3)在对称轴中心同时切割双链,产生平 末端或称钝性末端(blunt end),如SmaI:
5' …CCC▼GGG…3' 3' …GGG▲CCC…5' Sma I 5' …CCC 3' …GGG GGG…3' CCC… 5'
2. 限制酶HindIII的发现
Smith 等于1970年首次从流感嗜血杆菌(H. influenzae)中发现并分离到HindIII限制酶。
3. SV40 限制图谱和转录图谱的绘制
Nathans(1971年)用HindIII绘制SV40的 限制酶谱。
二、限制性内切核酸酶的类型
已经鉴定出的限制性内切核酸酶可分为4种 不同类型: Ⅰ型酶:能识别专一的核苷酸序列,并在 识别位点附近的核苷酸切割DNA双链,但 切割序列是随即的,无专一性。在基因工 程中应用不大。 Ⅱ型酶:能识别专一的核苷酸序列,并在 识别序列的固定位置切割双链DNA分子, 是基因工程中最常用的工具酶。
酶切反应注意事项 价格昂贵 决不能用水稀释,以免变性失活。 预先加入除酶以外的所有其他试剂。 取酶立即放于冰上。分装小份避免反复冻融。 使用无菌的新吸头。 少加水,使体积最小,• 保证酶液体积不超过总 但 体积的10%,否则酶液中的甘油会抑制酶活性。
2.2 DNA连接酶
一、DNA连接酶的发现 DNA连接酶(DNA ligase)是能催化双链 DNA片段靠在一起的3’羟基末端与5’端磷酸 基因末端之间形成磷酸二酯键,使两末端连 接在一起。—―分子黏合剂”
2hr 20hr 0 0 0 0 10% 75% >90 >90
EcoRI
GGAATTCC
6.反应缓冲液
pH范围: 7.0~7.6
缓冲液组份: Tris(三羟甲基氨基甲烷)、 NaCl、 MgCl2、二硫苏糖醇(DTT)或β巯基乙醇 有些酶需要BSA,以防止酶在低浓度蛋白 质溶液中变性。
7.酶的星号活性(star activity) 当条件改变时,许多酶的识别位点会改变, 导致识别与切割序列的非特异性,称为星号 活性。 EcoRⅠ正常情况下,其靶子序列为GAATTC, 但若缓冲液pH>8、盐离子浓度低于50mm和 甘油超过5%(V/V),可在NAATTN处切割, 以EcoRI*表示
六、限制内切核酸酶对DNA的消化作用
1.DNA分子的双酶切消化
可以先后分别在不同的反应体系中进行:
先用需要低盐离子浓度的酶切割,再调节 盐离子浓度,加入另一种酶切割。 先用最适反应温度较低的酶切割,升温后 再加入第二种酶。
2.DNA分子的完全酶切消化和部分酶切消化
完全酶切消化:内切酶对DNA序列中全部 的识别序列进行切割。 部位酶切:内切酶对DNA序列中所有的识 别序列进行不完全切割。
Bam HⅠ GGATCC CCTAGG Bg lⅡ AGATCT TCTAGA
G + GATCC CCTAG G
GATCT A + A TCTAG
五、影响限制内切核酸酶活性的因素
1.酶的纯度
2.DNA样品的纯度
高纯度DNA是必需的
样品如果残留氯仿、苯酚、乙醇、EDTA、 SDS等物质,或蛋白质、多糖等都可能影响 内切酶的反应活性。
DNA聚合酶Ⅰ的
重要用途是用于缺
口平移法制备核酸
的杂交探针。
Ⅱ. Klenow片段 Klenow片段具有DNA聚合酶Ⅰ的聚合酶活性 和3’-5’外切酶活性 ① 补平 3’ 凹端 DNA 。使用单纯的 DNA 合成活性。注意要加足够的 dNTP ;如果 使用带标记的 dNTP ,则可对 DNA 进行 末端标记。
(二)切割位点的特异性 1.DNA分子酶切片段的末端 切割:水解双链DNA中的磷酸二酯键,产 生的DNA片段5’端为磷酸基,3’ 端为羟基。
酶切后产生的片段末端有黏性末端和平末 端等形式。 黏性末端:DNA分子在限制性内切酶的作 用下形成的具有互补碱基的单链延伸形成 的末端结构。它们能够通过互补碱基间的 配对而重新连接起来。
三、DNA连接酶(ligase)的作用机理
四、影响连接反应的因素
1.反应温度: 黏性末端:4~15℃比较合适 平末端:10~20℃ 2.连接酶浓度: 平末端连接所需酶量较多 黏性末端连接所需酶量较少。
3.ATP浓度
一般情况下ATP最适的终浓度为0.5mmol/L。
(1)在对称轴两侧相对位点分别切割一条链。 靠近5’ 端切割产生 5' 端突出的粘性末端,如 EcoRI :
5' …G▼AATT C…3' 3' …C TTAA▲G…5' EcoR I 5’AATTC…3' 5' …G 3' …CTTAA 5’ G… 5'
(2)靠近 3' 端切割产生3' 端突出的粘性 末端。如Pst I:
Ⅲ型酶:有专一的识别序列,但不是对称 的回文序列,在识别序列旁边的位置上切 割双链。切割后产生的DNA片段具有各种 单链末端。
Ⅳ型酶:新鉴定出的一种酶,目前无应用。
三、限制性内切酶的命名
命名原则: 属——种——株——分离序号 例如:EcoRⅠ 属:Escherichia 种:coli 株:R 分离序号:Ⅰ
分为两类: 依赖于DNA的DNA聚合酶 依赖于RNA的DNA聚合酶-反转录酶
一、DNA聚合酶
Ⅰ. E.coli DNA聚合酶I(polI)-Kornberg酶
1)具两种外切酶活性和DNA聚合酶活性,在蛋白 酶作用下,该酶分裂成两个大小不同的片段,其中 小片段具5’→3’外切酶活性,大片段具3’→5’外切 酶活性(大片段亦称为Klenow片段, polIK) 2) 聚合酶活性,5’→3’, 要求3’-OH引物和模板 DNA,其延续性受反应条件的影响 3) 外切酶活性:具3’→5’和5’→3’外切酶活性
依据反应时所需能量辅因子的不同分为两类:
依赖ATP的DNA连接酶 依赖NAD+的DNA连接酶
依据其来源可分为三类:
T4噬菌体DNA连接酶、 大肠杆菌DNA连接酶 热稳定DNA连接酶。
二、DNA连接酶的特点
连接反应需要一条DNA链游离的3’-OH,另 一条链5’末端具有磷酸基团。 连接反应中需要ATP或NAD+和Mg2+为辅因 子和激活因子。 DNA连接酶并不能够连接两条单链的DNA 分子! DNA连接酶只能封闭双螺旋DNA骨架上的 缺口(nick),不能封闭失去核苷酸所形成 的裂口(gap)
有些限制性内切酶的识别序列中,某一位或 二位碱基并非严格专一,但不影响其作用位 点,只是增加酶的识别和作用频率。如 HindⅡ识别序列为--GTYRAC--,其中Y表 示C或T,R表示A或G。 有的酶识别6个以上的核苷酸序列,称为稀 有限制性内切酶,如NotⅠ,识别序列为 (GCGGCCGC)
①5’-3’聚合酶活性
条件 :
• Mg2+
•dNTP • 3’-OH
②3’- 5’外切酶活性
沿3’ 5方向识别和切除不配对的DNA生长 链末端的核苷酸。 在体内DNA复制时起校对作用,保证了 DNA复制的真实性,从而降低突变率。
③5’ - 3’外切酶活性
可降解DNA:RNA杂合体中的RNA分子 带切除的核酸分子必须具有5’端游离的磷酸 基团; 核酸分子被切除前位于已配对的DNA双螺旋 区段上; 被切除的可以是脱氧核苷酸,也可以是非脱 氧核苷酸
4.酶切反应的温度
大多数限制性内切酶的最适反应温度为37℃。
表2-3 部分限制性内切酶的最适反应温度 酶 ApaI ApyI BanI BclI BstEII 反应温度 30℃ 30℃ 50℃ 50℃ 60℃ 酶 MaeI MaeII MaeIII SmaI TaqI 反应温度 45℃ 50℃ 55℃ 25℃ 65℃
5.DNA分子的构型 某些限制酶,切割超螺旋的质粒DNA或病 毒DNA所需要的酶量,要比消化线性DNA 的高出许多倍。 还有些限制性核酸内切酶切割它们自己的 处于不同部位的限制位点,其效率也有明 显的差别。 大多数内切酶对只含识别序列的寡聚核苷 酸不具有催化活性。
HindIII
CAAGCTTG CCAAGCTTGG CCCAAGCTTGGG
命名 HindⅢ
属 种 株
Haemophilus influenzae d株 流感嗜血杆菌d株的第三种酶
序
第一个字母取自产生该酶的细菌属名,用大写; 第二、第三个字母是该细菌的种名,用小写; 第四个字母代表株; 用罗马数字表示发现的先后次序。
四、Ⅱ型限制内切酶核酸酶的基本特性
(一)识别序列的特异性
大多数Ⅱ型限制内切酶的识别序列长度为 4~6bp,具有双重旋转对称结构的回文序列 (palindromic sequence)。
BamHⅠ: GGATCC CCTAGG
如果识别序列由5对碱基组成,其对称轴为 中间的一对碱基,如MaeⅢ的识别序列为:
--GTNAC---CANTC-其中,N为任意碱基。
8.限制性内切酶反应的终止 消化DNA样品后,在进一步处理DNA样品 前往往需要钝化内切酶的活性。 钝化大多数限制性内切酶的方法是65℃温 浴5分钟。 但有些酶,如BamH I和HaeⅡ具备对热稳 定性,则需加入终止反应液(加入0.5mol/ L的EDTA使之在溶液中的终浓度达到 10mmol/L 以螯合Mg2+)。