动物个体发育模式
生物的个体发育

生物的个体发育生物的个体发育是指一个生物从单细胞状态经过生长、分化、发育等过程,最终形成成熟的个体的过程。
本文将从受精卵开始,介绍生物的个体发育的各个阶段及其特征。
受精和胚胎阶段生物的个体发育始于受精卵。
在受精卵中,精细胞与卵细胞相互结合,形成受精卵。
随着细胞的分裂和增殖,受精卵逐渐变成一小团细胞,这就是胚胎。
胚胎阶段是生物个体发育的前期,此时,胚胎不断分裂,细胞数量逐渐增多。
在这个过程中,细胞开始分化,形成不同的细胞类型,并逐渐组织成为不同的器官和组织。
胚胎还会进行一系列生理和行为变化,例如发育某些特殊的器官,如鳃、尾巴等。
胚胎发展的每一个阶段都有明显的特征,下面介绍几个常见的胚胎发育阶段:卵裂期受精卵形成之后,最初的几个细胞分裂后,核易位形成全细胞质膜,形成的细胞称为卵紊,这是胚胎发育的第一阶段,称为卵裂期。
囊胚期卵紊再分裂成为四个同等大小的细胞,这四个细胞继续分裂形成八个细胞,经过多次分裂,细胞数目逐渐增多,而每个细胞体积却逐渐缩小,直到形成囊胚阶段。
此时,胚胎就呈现出一个循环的、充满液体的囊状体。
原肠胚期在囊胚期之后,胚胎会通过一系列的细胞分化和移动形成一个植物或动物的基本组织结构。
在人类发育中,胚胎在发育到第三周时形成了原肠胚,即三个胚层的体系。
其中最外层是外胚层,最内层是内胚层,中间的一层是中胚层。
胚胎和成体阶段在经过不断的分化和增殖后,胚胎逐渐成为一个有机体,它的发展进入胚胎和成体阶段。
这个过程中,颜色、形状、大小、器官、组织等方面都会发生巨大变化,直到生物完全发育成熟。
在这个过程中,生物也会经历不同的发育阶段,例如人类就会分别经历早期胚胎期、胚胎期、胎儿期、婴儿期、儿童期、青少年期、成人期等不同的阶段。
总结总之,生物的个体发育是一个复杂的过程,需要经历许多步骤和阶段。
在这个过程中,每个阶段都有其特有的特点,生命体会在这个过程中慢慢长大、发育、进化。
我们应该认真学习、了解生物的发育过程,以更好地保护和利用生物资源,同时也推动生物学及其相关学科的发展。
多细胞动物的胚胎发育

一、个体发育和系统发育——系统发育
系统发育也可指 一个类群(如某个科、 属、种)的发生和发 展历史。
例如 :马的系统发生:经 历了六千万年的演变:
5
二、多细胞动物胚胎发育的一般规律
受精卵 囊胚
卵裂
原肠胚
中胚层和体腔形成
神经胚
幼体
6
二、多细胞动物胚胎发育的一般规律 1、卵裂期 1)卵的结构 植物极:卵黄多
囊胚
外胚层
原肠胚
内胚层
消化道的上皮
表皮及其 附属结构
(毛发、指 甲)
脑和神经系 统(感受器)
胚胎肠道 腺体(肝和脾)
其他:尿道和 呼吸道的上皮
膀胱的上皮
(气管、支气管、 肺上皮)
脊索
真皮(皮肤) 的内层
中胚层
循环系统(心脏、
血管淋巴系统等)
肌肉(骨骼肌、平滑肌)
内脏器官的外膜
排泄系统(肾、输尿管)
生殖系统(睾丸、卵巢、 输卵管、子宫)
蛙受精卵的特点: 动物半球:卵黄少、比重小、颜色深 植物半球:卵黄多、比重大、颜色浅
卵裂:辐射卵裂 原肠胚特点:从新月区的中心开始 胚的发育:受精卵经细胞分裂、组织分化、器官形
成,形成幼体的过程 胚后发育:幼体从卵膜里孵化出来或从母体里生出
来,并发育成成体(性成熟的个体)的过程
34
二、两栖动物的胚胎发育——蛙受精卵分裂的过程
动物 半球 囊胚腔 植物 半球
外胚层
中胚层 原肠腔 内胚层
37
动物的个体发育总结
受精卵
卵裂
胚
囊胚 (具囊胚腔)
的
动物半球细胞外包
发
植物半球细胞内陷
育
原肠胚 (具原肠腔、三个胚
高等动物的个体发育详解

体 发 育
分化
组织、器官分化 系统形成
胚
幼体 (从卵膜中孵化或母体生出)
后
温度等外界因素
发
育
成体
巩固练习:
1、能正确表示高等动物个体发育顺序的是(B ) A、卵 幼体 成体 B、受精卵 幼体 成体 C、受精卵 卵裂 囊胚 原肠胚 组织器官的分化 D、卵 卵裂 囊胚 原肠胚 组织器官的分化
2、人体的皮肤来源于( D ) A、内胚层 B、外胚层 C、中胚层 D、外胚层和中胚层
试管婴儿技术是体外受精、胚胎 移植等人工助孕技术的俗称,是一 项结合胚胎学、内分泌、遗传学以 及显微操作的综合技术。它是将精 子和卵子置于体外利用各种技术使 卵子受精,培养几天后移入子宫, 使女性受孕生子。
这是1979年9月7日,世界上第 一个试管婴儿路易丝·布朗和她的父 母一起在芝加哥的电视节目中和观 众见面。
原肠胚
组织 器官 系统的分化与形成
外胚层 中胚层 内胚层
表皮及其附属结构 神经系统和感觉器官
脊索、真皮、肌肉 内脏器官的外膜 排泄系统、生殖系统、循环系统
消化道上皮、呼吸道上皮 腺体(如肝、胰等)
鸡的胚胎
人类胚胎第8周时在子宫内的照片
爬行动物、鸟类和哺乳类在胚胎发育早期,从胚胎四周的表 面开始形成围绕胚胎的胚膜。胚膜的内层称为羊膜。由羊膜 所包围的腔称羊膜腔。腔内充满羊水,胚胎悬浮其中。
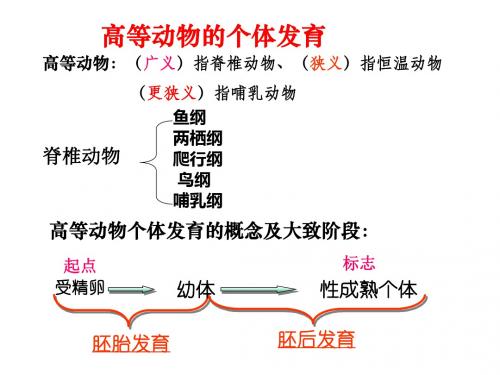
高等动物的个体发育
新野一高
王怡娜
动物个体发育阶段
高等动物的个体发育可以分为:胚胎发育和胚后发育
胚胎发育
受精卵发育为幼体
胚后发育
幼体从卵膜内孵化出来→成体 (如卵生动物)
幼体从母体内生出来→成体
(如胎生或卵胎生动物)
高等动物的个体发育过程

一、高等动物的个体发育过程1.起点::受精卵。
2.过程:受精卵幼体性成熟的个体。
3.概念(1)胚胎发育:受精卵发育成为幼体的过程。
(2)胚后发育:幼体从卵膜孵化出来或从母体生出来后,发育成为性成熟个体的过程。
皮肤和胃分别是由哪些胚层发育而成的?二、胚胎发育1.过程受精卵卵裂概念:受精卵早期的细胞分裂囊胚原肠胚幼体:已经经过细胞分化,形成了具有组织、器官的完整个体2.爬行类、鸟类和哺乳类动物个体发育的特点及意义(1)特点:胚胎发育早期,具有羊膜和羊水。
①羊膜:指胚胎发育早期,从胚胎四周的表面开始,形成的围绕胚胎的胚膜的内层。
②羊水:羊膜内的液体。
(2)意义①保证了胚胎发育所需的水环境。
②具有防震和保护作用。
使动物增强了对陆地环境的适应能力。
【思考·提示】皮肤由外胚层和中胚层发育而成,胃由中胚层和内胚层发育而成。
三、胚后发育核心知识一、胚胎发育1.过程受精卵:卵黄富含营养物质,卵裂:受精卵早期的有丝分裂多细胞球状胚,继续细胞分裂,内部形成空腔囊胚:里面的空腔称囊胚腔,表层细胞向内运动形成内层细胞;进一步在内外细胞层之间形成一个新细胞层成都家教、绵阳家教原肠胚:有外、中、内三个胚层;中间具有原肠腔和缩小的囊胚腔,细胞进一步分化,形成组织、器官、系统幼体2.营养供给哺乳动物胚胎发育早期所需营养物质由卵黄提供,后期由母体通过胎盘提供。
其他动物的胚胎发育所需营养均由卵黄提供。
【特别提醒】(1)囊胚与胚囊不同。
囊胚是动物胚胎发育过程中的一个特定时期,此时的胚胎是一个内部有囊胚腔的球状胚。
而胚囊是被子植物雌蕊中属于子房中胚珠的组成部分。
(2)胚胎发育各期的细胞,越早期的细胞分化程度越低,其全能性越高,受精卵的全能性最高。
1.能正确表示高等动物个体发育顺序的是( )A.卵→幼体→成体B.受精卵→卵裂→囊胚→原肠胚→组织、器官分化C.受精卵→幼体→成体D.卵→卵裂→囊胚→原肠胚→组织、器官分化解析:选C。
12-13动物的繁殖和个体发育

植物极
蛙囊胚形成
两栖类 囊胚壁 多层细胞 内细胞团 无
哺乳类 单层细胞(滋养层) 有,将分化为胚盘
几种囊胚的比较
受精卵(第一天)
雌原核和雄 原核靠近
二核融合开始分裂(卵裂)
内细胞团
滋养层
胚泡腔
2细胞期
4细胞期 (第二天)
桑椹胚 (第三天)
囊胚或胚泡 (第四天)
受精、卵裂、胚泡形成与胚泡逐渐植入子宫内壁 (植入大约在第5-6天开始,到第11-12天结束)
动物的繁殖和个体发育
蒲淑萍
生物繁殖的基本类型
• 无性生殖
裂殖(二分裂生殖):细菌、原生动物 芽殖(出芽):酵母菌、水螅 断裂生殖(再生):海绵、涡虫、海星、珊瑚、植物扦插 单性生殖(孤雌生殖):卵未经受精而直接发育成雄性个体 (蜜蜂、蚂蚁)或雌性个体(轮虫、蚜虫、水蚤) 同卵双生:人类的同卵双胞胎
●受精过程 ①顶体反应 ②精卵质膜融合与雌雄原核形成 ③皮层反应与透明带反应 问题:如何避免多精受精? = 为何只有一个精子能进入卵子? = 卵子如何阻止其他精子再进入的?
●受精方式
①体外受精:雌雄个体同时释放出精子和卵→在水中 完成受精,如珊瑚、硬骨鱼类、两栖类 ②体內受精:雌雄先行交配→在雌体內完成受精,如 甲壳类、昆虫、软骨鱼类、爬行类、鸟类、哺乳类
3、成熟期
次级卵 母细胞 第一 极体
减数分裂Ⅱ, 排卵时停留在 中期Ⅱ 受精后,完成 减数分裂Ⅱ
初级卵 母细胞
性激素 恢复完成 减数分裂I
卵细胞、 第二极体
(n)
减数分裂Ⅱ
2个第 二极体
(n)
卵 细 胞 的 形 成 过 程
卵子减数分裂时间比精子更长,更易发生和积累染色体变异。
动物的发育模式

(五)动物体腔的发生方式不同 1.无体腔动物:中胚层发育中不与其它 胚层联合或者自身不发生腔裂的动物。 如扁形动物等。 2.假体腔动物:囊胚腔残留到成体期形 成体腔,位于中胚层和内胚层之间。
如线虫动物、线形动物等。 3.真体腔动物:体腔由中胚层内部的腔
裂和扩展而成。 如软体动物、环节动物、节肢动
物和所有的脊索动物等。
原肠胚
原肠胚
原肠胚
原肠胚
消化管ቤተ መጻሕፍቲ ባይዱ形成
囊胚 受精卵
中央细 胞凋亡
细胞迁 移形成 中胚层
形成神经系统 器官、系统发生
图:文昌鱼的胚胎发育
神经胚 幼虫
幼虫 幼虫
(二)不同物种之间的胚胎发育有很大差别
1.肛肠动物:棒螅没有囊胚阶段。 钵水母有囊胚阶段。
2.棘皮动物:海胆没有神经胚阶段。 3.高等脊椎动物鸟类和哺乳动物:具有
第一节胚胎学对动物发育模式的确定和阶段划分第二节模式动物第一节胚胎学对动物发育模式的确定和阶段划分一动物的生活周期一世代交替重点在多细胞生物周期性的生活史中存在有双倍染色体和单倍染色体两种不同细胞构成的阶段并且它们之间相互更替出现
第四章 动物的发育模式
第一节 胚胎学对动物发育模式的确定和 阶段划分
第二节 模式动物
海绵
水螅
海胆及其贝壳
2.辐射对称型:肛肠动物水螅、棘皮动物海胆
3.两侧对称型:人类等。
(二)胚胎的卵裂方式不同
(完全卵裂和不完全卵裂)
辐射型(海胆)
螺旋型 软体动物、环节动物
背 前
1. 完全卵裂
旋型
背 前
、环节动物
的4
种类型
腹
后
对称型(背囊动物)
旋
第二章多细胞动物的胚胎发育_动物生物学

一、动物的个体发育和系统发育的概念1、个体发育(ontogeny)是指多细胞动物体从受精卵开始,经过细胞分裂、组织分化、器官形成,直到性成熟的全过程。
高等动物的个体发育包括胚胎发育和胚后发育两个阶段。
2、系统发育(phylogeny)也称系统发展,是与个体发育相对而言的,它是指某一个类群的形成和发展过程。
二、卵细胞的极性、卵裂的形式(一)卵细胞的极性卵细胞的极性(图2-1-1 卵细胞的极性)是指卵细胞的内部结构是非均向性的即细胞核的位置和细胞质分布的不对称性。
通常将卵黄多的一端称为植物极,另一端称为动物极。
(二)卵裂的形式卵裂(cleavage)(图2-1-2 卵裂)即是受精卵进行分裂。
根据不同类动物卵内卵黄多少及分布情况的不同,将受精卵的卵裂分为:1、完全卵裂:多见于少黄卵,整个卵细胞都进行分裂。
完全卵裂又包括等裂和不等裂。
①等裂:是指卵黄少、分布均匀、形成的分裂球大小相等,如文昌鱼、海胆等。
②不等裂:是指卵黄分布不均匀,形成的分裂球大小不等,如多孔动物、蛙类等。
2、不完全卵裂:多见于多黄卵。
由于卵黄多,分裂受阻,受精卵只在不含卵黄的部位进行分裂。
不完全卵裂又包括盘裂和表裂两种:①盘裂:指分裂区只限于胚盘处的分裂。
如乌贼、鸡卵等。
这是由于卵黄物质多,细胞核和细胞质集中于卵一端的缘故。
②表面卵裂:分裂区只限于卵的表面的分裂。
如昆虫卵。
这是由于大量卵黄集中在卵的中央所致(图2-1-3 龙虾的表面卵裂)。
(二)卵裂的形式卵裂(cleavage)(图2-1-2 卵裂)即是受精卵进行分裂。
根据不同类动物卵内卵黄多少及分布情况的不同,将受精卵的卵裂分为:1、完全卵裂:多见于少黄卵,整个卵细胞都进行分裂。
完全卵裂又包括等裂和不等裂。
①等裂:是指卵黄少、分布均匀、形成的分裂球大小相等,如文昌鱼、海胆等。
②不等裂:是指卵黄分布不均匀,形成的分裂球大小不等,如多孔动物、蛙类等。
2、不完全卵裂:多见于多黄卵。
由于卵黄多,分裂受阻,受精卵只在不含卵黄的部位进行分裂。
动物个体发育 PPT课件 人教版

心脏两心房 一心室
返回
受精卵的特点:
颜 色深
动物极 卵 黄 少
比 重小
朝向上端
颜色浅
卵黄多
植物极
比重大
朝向下端
受精卵
受精卵
受精卵
第一次卵裂
第二次卵裂
第三次卵裂
囊胚
囊胚腔
囊胚
பைடு நூலகம்重复
植物极细胞內陷
原肠腔
原肠胚
原肠胚
囊胚腔
原肠腔
原肠胚
原肠胚
•
17、一个人只要强烈地坚持不懈地追求,他就能达到目的。你在希望中享受到的乐趣,比将来实际享受的乐趣要大得多。
•
18、无论是对事还是对人,我们只需要做好自己的本分,不与过多人建立亲密的关系,也不要因为关系亲密便掏心掏肺,切莫交浅言深,应适可而止。
•
19、大家常说一句话,认真你就输了,可是不认真的话,这辈子你就废了,自己的人生都不认真面对的话,那谁要认真对待你。
高中生物课件 管炜平
引入新课 胚的发育 板书总结1 2
高等动物 的个体发育
动画
胚后发育
思考与练
制作人: 广丰实验中学高二年级
习 管炜平
雄性个体
雌性个体
减数
减数
分裂
分裂
精子
卵子
生
殖
受精作用
受精卵
细胞 组织
分裂 分化
个
器形
体
官成
发 育
新个体
个体发育:
受精卵经过细胞分裂、组织 分化、器官形成而发育成新个 体的过程
有丝分裂:
分裂产生的子细胞经过生长再分裂
返回
1. 除哪项外,其余都是原肠胚的特点[ B ]
动物的繁殖和个体发育

从囊胚到原肠胚:海星
囊胚腔
原肠腔
外胚层 中胚层 内胚层 胚孔
囊胚
原肠胚
囊胚中期
尾鳍出现期
第二节 动物的个体发育
三、胚后期 包括生长和发育两个过程 1. 变态发育或间接发育:动物的胚后期分两个
阶段,即幼体(虫)期和成体(虫)期。幼体在 形态和生活习性上与成体明显不同,幼体(虫) 必须经过复杂的变化过程(即变态),才能发育 为成体(虫)。如昆虫和青蛙等。 2.无变态发育或直接发育:胚后发育没有幼体 (虫)期之分。其早期发育阶段在形态和生活习 性上与成年期基本相同。
第一节 动物的繁殖
3.其它繁殖方式:特殊繁殖方式,包括: (1)接合生殖,如草履虫; (2)孤雌生殖(有性个体进行的无性繁殖),如轮 虫和水蚤; (3)幼体生殖(尚未达到性成熟的个体进行的无性 繁殖) ,如三代虫; (4)世代交替(有性生殖和无性生殖有规律地交替 进行),如薮枝螅。
动物的繁殖和个体发育
点斑隼蟌
第一节 动物的繁殖
一、繁殖的意义:种族延续和进化发展 二、繁殖的方式: 1. 无性繁殖:原始的繁殖方式,遗传物质来
源于一个亲体,其特点是繁殖速度快,但种 族进化发展慢。主要有三种 :(1)分裂 (有二分体分裂即二分裂,如眼虫和多分体 分裂即复分裂,如疟原虫);(2)断裂,如 涡虫;(3)出芽,如水螅和海鞘等。
第二节 动物的个发育
一、胚前期 胚前期是指精子和卵子的形成期,是动物生命的预 备阶段。 1. 精子的形成包括:增殖期、生长期、成熟期和 变形期四个阶段,1――4 2. 卵子的形成包括:增殖期、生长期和成熟期三 个阶段,1――1
第二节 动物的个体发育
二、胚胎期 胚胎期以精卵结合为起点,至幼体从母体娩出或从 卵中孵出结束。 1. 受精:有体内受精或体外受精两种 2.胚胎发育:通常包括:卵裂、囊胚形成、原肠胚 形成、中胚层及体腔形成等几个阶段。
动物的成长与发育

动物的成长与发育
动物的成长与发育是生物学中一项重要的研究内容。
它涉及动物从胚胎发育到成熟个体的各个阶段和过程。
动物的成长过程通常可以分为以下几个阶段:
1. 胚胎期:动物从受精卵开始发育,逐渐形成胚胎。
在这个阶段,细胞不断分裂、增殖,形成各种组织和器官的前体。
2. 幼体期:胚胎发育完成后,动物进入幼体期。
在这个阶段,动物逐渐长大,体型增大。
它们开始独立获取食物,并适应环境中的各种变化。
3. 成体期:幼体期过后,动物进入成体期。
在这个阶段,动物已经完全成长发育,体型达到成年大小,并且具备繁殖能力。
它们会进行繁殖、生存以及与其他个体的交流和互动。
动物的成长和发育过程受到多种因素的影响,包括遗传因素、环境因素、营养摄取等。
不同种类的动物在成长发育过程中存在差异,有些动物在短时间内即可成熟,而有些动物则需要较长时间。
理解动物的成长与发育对于生物学研究以及动物保护和养殖都具有重要意义。
通过深入研究动物的成长与发育规律,可以更好地了解动物的生态特征,为它们的保护提供科学依据。
同时,对于动物的养殖来说,了解其成长和发育过程可以帮助提高养殖效率和管理质量。
总而言之,动物的成长与发育是一个复杂而有趣的过程,它关系到动物的生存和繁衍。
对于科学研究、生物学教学以及动物保护和养殖都具有重要意义。
高等动物的个体发育
高考资源网,您身边的高考专家!
精子和卵细胞在水中进行受精,形成受精卵。
高考资源网,您身边的高考专家!
蛙胚的发育过程(一)
1.受精卵
卵轴
动物极
位置: 上端 卵黄少、
动物半球 特点: 比重小、
颜色深
作用:吸光热,提供温度条件
对于个体发育有何意义
位置: 下端
植物半球
卵黄多、
植物极 特点: 比重大、
蛙
胚
后
尾 四肢 呼吸器官 生活环境
发
蝌蚪 有 无
鳃
水中
的
幼蛙 无 有 肺、皮肤 陆地、水中
育
过 【探索】 蝌蚪具备的特征跟哪类生物最相近,
程 说明了两类生物之间有怎样的关系?
鱼类、两栖类是由古代鱼类进化而来的。 高考资源网,您身边的高考专家!
高考资源网,您身边的高考专家!
1、下列不属于胚后发育的是 ( A ) A、鱼的受精卵发育成鱼苗 B、蝌蚪从卵膜里孵化出来后发育成青蛙 C、家蚕发育成蚕蛾 D、小鸡破壳而出后发育成鸡
机 物
种类: 增多
6、结合本节所学内容,思考在由受 精卵发育成原肠胚的过程中,
所有细胞DNA总量变化曲线是 A 每个细胞中染色体数目的变化曲线是 B 所有细胞中有机物总量的变化曲线是 C
高考资源网,您身边的高考专家!
种类 数量
时间
A
B C
时 间
4、下图表示蛙的受精卵发育至囊胚过程中,DNA总 量、每个细胞体积、所有细胞体积之和、有机物总 量的变化趋势(横坐标为发育时间)。其中正确的是
( A)
A.①②
B.①③ C.②④ 高考资源网,您身边的高考专家!
D.③④
第三章 动物的繁殖与个体发育

第三章动物的繁殖与个体发育第一节繁殖的概念和方式一、繁殖的概念:动物产生后代的现象称之为繁殖。
是种群的延续。
二、繁殖的方式无性繁殖:不经生殖细胞的结合,由亲体直接产生子代的繁殖方式。
多见低等动物。
1、分裂生殖复分裂法(裂体生殖):2、出芽生殖3、孢子生殖:孢子虫所特有。
母体产生许多孢子,不经结合直接形成新个体。
4、再生(断裂生殖)有性繁殖:是由雌雄细胞结合而产生新个体的生殖方式。
1、配子生殖:性细胞(配子)结合产生新个体同配生殖:两个结合的配子,形态大小相同,仅在生理上有区别的。
卵生:母体产出的是卵,胚胎在体外发育,营养来自卵黄异配生殖胎生:母体产出的是幼体,体内发育,营养来自母体卵胎生:母体产出的是幼体,体内发育,营养来自卵黄2、接合生殖:纤毛虫所特有,接合时两个体以口沟部分相接合,表膜溶解,细胞质通连,小胞核交换,相互融合。
两虫体分开后,各自分裂增殖。
几种特殊繁殖方式:孤雌生殖(单性生殖):即雌性所产生的卵,不经受精能直接发育成新个体。
如轮虫、蚜虫等。
在蜜蜂中,蜂王产的不受精卵发育成雄蜂,也是一种孤雌生殖。
幼体生殖:动物个体未成熟期或幼体阶段,即能进行繁殖,如昆虫中的瘿蝇等。
多胚生殖:受精卵形成多个胚胎新个体世代交替:无性生殖和有性生殖有规律性交替出现。
如腔肠动物中的某些种。
第二节动物的繁殖与个体发育发育:在动物的生活史中或演化过程中,形态、结构和机能由简单到复杂的变化过程叫发育。
因此,发育可分为个体发育和系统发育。
个体发育:胚前期、胚胎发育(受精和受精卵、卵裂、囊胚、原肠胚、胚层及体腔的形成、三胚层分化)和胚后发育(直接和间接发育)一、胚前发育:此期发育始于精、卵细胞的产生,终于其成熟。
卵具极性动物性极—动物卵细胞内核、质所在,细胞质较多、卵黄较少的一端,比重小、位于上方,卵裂时分裂快的一极。
植物性极—动物卵细胞内比重大、卵黄多、位于下方的一极,卵裂时分裂慢或不能分裂。
卵的类型少黄卵(均黄卵):哺乳动物、文昌鱼等多黄卵偏黄卵:黄卵偏植物极。
动物的生长发育从幼小到成长

动物的生长发育从幼小到成长动物的生长发育是一个复杂而持续的过程,从幼小到成长,动物经历了多个阶段的变化和发展。
这个过程涉及到生理、形态和行为等方面的变化,不同动物在生长发育方面也存在一些共同的特点和差异。
本文将就动物的生长发育过程进行探讨。
一、胚胎期胚胎期是动物生长发育的第一个阶段,它从受精卵形成开始,一直持续到胚胎形成。
在这个阶段,动物的细胞开始分化和定位,逐渐形成不同的组织和器官。
胚胎期对于动物的后续发育具有重要的影响,一旦出现异常,可能导致发育缺陷或畸形。
二、幼年期幼年期是动物生长发育的第二个阶段,也是从幼崽出生或孵化开始,到达性成熟前的阶段。
在这个阶段,动物处于生长的黄金时期,身体逐渐长大,各种器官和系统逐渐完善。
幼年期也是动物学习和适应环境的关键时期,很多动物在这个阶段需要依靠父母或其他成年个体的照顾和保护。
三、青少年期青少年期是动物生长发育的第三个阶段,也是从幼年期到达性成熟前的阶段。
在这个阶段,动物身体继续增长,各个器官和系统进一步发育成熟。
青少年期是动物个体形态发生明显变化的阶段,例如鸟类的羽毛颜色变化、哺乳动物的骨骼增长等。
同时,动物的行为也逐渐呈现出成年个体的特征。
四、成年期成年期是动物生长发育的最后一个阶段,也是动物生命中最长的一个阶段。
在这个阶段,动物的身体发育基本停止,各个器官和系统已经完全发育成熟。
成年期的动物已经具备了繁殖能力,开始繁衍后代。
不同种类的动物成年期的长度和特征也存在很大的差异,例如某些动物的成年期只持续几个月,而象、鲸等大型动物的成年期可能长达几十年甚至更长。
总结起来,动物的生长发育从幼小到成长经历了胚胎期、幼年期、青少年期和成年期等不同阶段。
每个阶段都有着特定的生理、形态和行为变化,是动物适应环境并生存繁衍的关键时期。
对于了解动物的生态习性和进化过程有着重要的意义。
动物的生长发育是一个奇妙而复杂的过程,我们应当珍惜和保护好每一个动物个体的成长。
动物学资料 动物体的基本结构

端 细 胞 法
(裂)
体 腔 囊 法
(肠)
体节进一步分化为三部分: (1)生骨节(sclerotome),为体节的内侧部分,将来 形成脊索鞘和神经管外面的结缔组织以及肌隔等; (2)生肌节(myotome),为体节中间部分,将来形成 肌节; (3)生皮节(dermatome),为体节的外侧部分,将来 形成表皮下面的结缔组织,即真皮。
1.神经管的形成:胚胎背面沿中线的外胚层细胞向下陷,形成神经
板(neural plate)。神经板两边的外胚层细胞首先翘起形成一对 纵褶,其后两边的纵褶逐渐靠拢,除前端形成神经孔的一小部分外, 完全愈合于中线。同时下陷到表皮内的神经板的两侧向上弯曲,最 后两边缘在背面闭合,形成背面有一缝隙的神经管(neural tube), 管中央的腔隙,称神经管腔(neurocoel),在前端以神经孔 (neuropore)和外界相通,到成体时,该孔关闭,成为嗅窝。胚 胎在这一时期神经管腔和原肠仍以神经肠管(neurenteric canal) 相通,以后神经肠管消失,神经管和原肠就互不相通了。此时期的 胚胎称为神经胚(neurula)。
动物体的基本结构
3.2.1 对称
二、辐射对称:通过其身体的中央轴可有 两个或两个以上切面把身体分割成相等的 两部分。
动物体的基本结构
三、两侧对称: 通过身体的中央轴,只有一 个切面将动物体分成左右相等的两部分。
两侧对称的出现在动物进化上的意义:
1)凡两侧对称的动物身体都有了明显的前后、背腹、左右之
动功能;C.脊椎动物的内骨骼:主要功能是支撑, 是运动系统一部分,防护功能由皮肤承担。
•
分节的意义
• 动物身体的分节是生理分工的开始,使得
各种器官位于一定的体节中,在比较高等 的动物中就形成了头、胸、腹的分部。
动物的发育

一、高等动物的个体发育类型高等动物的个体发育包括胚胎发育:发育成为幼体胚后发育:二、胚胎发育蛙胚的发育过程卵裂的结果:内含的空腔称、此时胚的外形为,囊胚腔逐渐,形成了新的腔为,由于极一端的细胞分裂较快,新产生的细胞便向方向推移,使得极一端的细胞向囊胚腔,使囊胚腔逐渐。
这些陷入的细胞不仅构成了胚胎的胚层,而且围成了一个新的腔,即腔。
接着细胞层之间分化出一个新的细胞层,即胚层。
此时期的胚胎具有、和三个胚层,叫做胚。
(3) 类、类和哺乳类等动物,在胚胎发育的早期,形成围绕胚胎的膜,其内层叫膜。
它呈囊状,里面的液体叫。
羊膜和羊水不仅保证了胚胎发育所需的环境,还具有和作用,增强了这些动物对环境的适应能力。
3.胚后发育。
爬行动物、鸟类和哺乳动物的幼体和成体差别较小,对这些动物来说,其胚后发育主要是指和生殖器官的,而有些动物的幼体和成体,在和上却有明显差异,如。
这类动物在发育的过程中的特点:和都要发生显著的变化,这些变化又是集中在内完成的。
1987年7月25日的深夜)1时47分,世界上第一例”试管婴儿”路易斯·布朗问世了,小布朗的产生过程大致如下:取母亲的卵细胞和父亲的精于在试管内完成受精,等受精卵在试管内发育至囊胚时,再移入母亲的子宫内,胚胎经过9个月的发育,小布朗问世了.试问小布朗的形成属于动物个体发育的哪个阶段?在试管内发育的是哪几个阶段?在母体子宫内发育的是哪几个阶段?2.没有交配的母鸡产下的一个鸡蛋是否是一个卵细胞?3.青蛙的幼体与成体有哪些差别?除了两栖动物外,还有哪些动物也具有变态发育的特点?试举例说明之.例1)下图表示蛙的受精卵寥育至囊胚过程中,DNA总量、每个细胞体积,所有细胞体积之和、有机物总量的化趋势(横坐标为发育时间).其中正确的是( )久①②B.①③C②④D.③④[例2]下列关于羊膜出现的说法,错误的是……………( )A.羊膜内的羊水为胚胎发育提供了水环境B羊膜内的羊水具有防震和保护作用.C.羊膜的出现使动物更加适应陆地环境D。
动物的繁殖与发育

动物的繁殖与发育动物的繁殖与发育是生物学中一个重要且广泛研究的领域。
通过繁殖和发育的过程,动物物种能够延续生命,保持种群数量的稳定,并适应环境的变化。
本文将介绍动物的繁殖与发育的基本概念、不同动物群体的繁殖与发育方式以及一些与繁殖和发育相关的特殊现象。
一、动物的繁殖与发育概述动物的繁殖与发育是指动物个体的生殖细胞(精子和卵子)在合适的生理和环境条件下结合并形成新的个体。
这个过程涉及到生殖细胞的产生、结合、受精、胚胎的发育、胚胎的成长和后续的个体发育等一系列的阶段。
动物个体的繁殖方式可以分为两大类:性别繁殖和无性繁殖。
性别繁殖是指动物个体通过交配和受精产生新的个体,其中雌性产生卵子,雄性产生精子。
而无性繁殖则是指个体直接通过无性生殖方式产生新的个体,不需要受精过程。
二、不同动物群体的繁殖与发育方式在动物界中,不同物种的繁殖与发育方式差异很大。
以下是一些常见动物群体的繁殖与发育方式的介绍:1. 爬行动物:爬行动物包括蜥蜴、蛇和龟鳖等。
大部分爬行动物采取卵生繁殖方式,即雌性动物产卵,然后卵在外界孵化,孵化出的幼体经过一段时间的发育变为成体。
2. 鸟类:鸟类是以卵生繁殖方式为主的动物群体。
雌鸟通过产卵的方式将卵留在巢中,雄鸟同样参与孵化过程。
一段时间后,卵孵化出幼鸟,幼鸟通过一系列的发育过程逐渐成长为成鸟。
3. 哺乳动物:哺乳动物的繁殖方式较为复杂。
雌性哺乳动物的卵子在体内受精,形成受精卵,然后嵌入子宫壁内继续发育,最终通过胎盘供给营养给胎儿。
一段时间后,胎儿发育成熟,通过分娩的方式出生。
4. 昆虫:昆虫是在动物界中数量众多的一个类群。
昆虫的繁殖方式多样,包括卵生、卵胎生和胎生等。
卵生昆虫通过产卵产生新的个体,卵孵化后产生幼虫,幼虫经过若干个体蜕变阶段最终成为成虫。
而卵胎生和胎生昆虫则是在体内孵化出幼虫或直接产生幼虫。
三、与繁殖与发育相关的特殊现象在动物界中,还存在一些与繁殖和发育相关的特殊现象,下面介绍两个典型的例子:1. 轮回生殖:某些无脊椎动物(如水螅、螺旋藻等)具有轮回生殖的特性。
高等动物的个体发育

高等动物的个体发育【自学导引】一、高等动物的个体发育类型:高等动物的个体发育包括二、胚胎发育1.胚的发育过程2.羊膜的出现及其意义某些高等动物,在胚胎发育的早期,从胚胎四周的表面开始形成了围绕胚胎的胚膜。
胚膜的层叫羊膜。
羊膜里面充满了液体,叫羊水。
羊膜和羊水不仅保证了胚胎发育所需的液体环境,还具有防震和保护作用。
三、胚后发育1.爬行类、鸟类、哺乳类的胚后发育(1)大致过程:幼体(幼体从卵膜孵化出来或从母体生出来)→成体(2)胚后发育主要指身体的长大和生殖器官的逐渐成熟。
2.青蛙的胚后发育过程:蝌蚪(幼体)→幼蛙→成蛙。
特点:变态发育——某些动物在胚后发育的过程中,形态结构和生活习性都要发生显著的变化,而且这些变化又是集中在短时期完成的。
【思考导学】1.1987年7月25日的深夜11时47分,世界上第一例“试管婴儿”路易斯·布朗问世了。
小布朗的产生过程大致如下:取母亲的卵细胞和父亲的精子在试管完成受精,等受精卵在试管发育至囊胚时,再移入母亲的子宫,胚胎经过9个月的发育,小布朗问世了。
试问小布朗的形成属于动物个体发育的哪个阶段?在试管发育的是哪几个阶段?在母体子宫发育的是哪几个阶段?答案:胚的发育;受精卵→囊胚;囊胚→原肠腔→幼体。
2.没有交配的母鸡产下的一个鸡蛋是否是一个卵细胞?答案:不是,卵细胞只包括胚盘、卵黄及卵黄膜,把一个鸡蛋看作是一个卵细胞,是不妥的。
3.青蛙的幼体与成体有哪些差别?除了两栖动物外,还有哪些动物也具有变态发育的特点?试举例说明之。
答案:青蛙的幼体是蝌蚪,蝌蚪有尾巴没有四肢,青蛙有四肢没有尾巴,蝌蚪用鳃呼吸,青蛙用肺呼吸,兼用皮肤呼吸,蝌蚪的心脏是一心房一心室,一条循环路线(无体循环和肺循环之分),青蛙的心脏是二心房一心室,二条循环路线(体循环和肺循环);青蛙可以到陆上生活,肉食性(即主要以昆虫等小型无脊椎动物为食),蝌蚪是植物食性的,以水中的藻类为食。
昆虫都具有变态发育的特点,如家蚕。
动物的个体发育

动物的个体发育动物的个体发育动物的个体发育问题1:动物个体发育的起点,终点及阶段起点:受精卵.终点:性成熟.可分为:胚胎发育和胚后发育两个阶段.其界限是幼体从卵膜内孵化出来或从母体生下来. 可表示为:受精卵(起点)胚胎发育幼体胚后发育成体(终点)问题2:动物胚胎发育要依次经历哪几个阶段(1)从受精卵开始首先经历卵裂.卵裂实质是有丝分裂,但与有丝分裂又有不同,即细胞数量越分越多,细胞和体积越来越小(只分裂不生长).(2)细胞经3次卵裂形成八细胞胚胎.此时开始出现细胞不均等现象.(细胞的大小,含营养物质的多少等)(3)然后由于营养的消耗,细胞分裂速度的快慢等原因,形成一个内有空腔的囊胚.形成原因:动物半球细胞外包和植物半球细胞内陷.问题3:原肠胚有什么特点A,有原肠腔:将来发育成消化道,是原肠胚形成的标志.B,有内,中,外三个胚层:内胚层来自内陷的植物半球细胞分化形成.外胚层由外包的动物半球细胞分化形成,中胚层中既有植物半球细胞,也有动物半球细胞.(所以高等动物的胚胎发育一般认为从原肠胚时期开始出现细胞分化,此后细胞的全能性程度逐渐减小).三个胚层将分别分化发育成动物的各种组织,形成各种器官和系统,进而形成一个完整的新个体.这一过程我们称为组织器官分化形成期.幼体再经胚后发育形成性成熟个体.高等动物的胚后发育通常有两种模式:直接发育和变态发育,其中变态发育的代表生物类群有:蛙,几乎所有的昆虫.表现出来.种系发育又制约着个体发育,或说个体又简单地,发育是一个过程,至少包括着性状的发育,个体发育和种系发育.性状发育是指的基因控制性状的表达过程;个体发育是从生到死的个体生命的全过程;种系发育是指的每一物种的系统发育,或是该物种的演变过程.这三者是互相联系的,性状的发育在个体发育过程中实现,例如某男性个体具有秃顶基因,秃顶性状也许要个体发育到35岁以后才渐渐表现出来.种系发育又制约着个体发育,或说个体又简单地,迅速地重演种系发育.。
高等动物的个体发育过程PPT课件

基因编辑技术还可能影响生物的进化过程,通过修改种群的基因频率,实现对生物种群的定向进化。
生物进化
基因编辑技术的应用
干细胞是一类具有自我更新和多向分化潜能的细胞,能够分化成各种组织和器官,为高等动物个体发育和再生医学研究提供了新的思路。
干细胞
干细胞治疗是一种新兴的治疗方法,通过将健康的干细胞移植到患者体内,替代或修复病变细胞,达到治疗疾病的目的。
口腔、食管、胃、小肠和大肠等器官的发育,使得动物能够消化和吸收食物中的营养。
01
呼吸系统的发育
随着肺的发育,高等动物能够进行有效的气体交换,支持生命活动。
02
循环系统的发育
心脏和血管系统的发育使得血液能够有效地在体内循环,输送氧气和营养物质。
生理机能的发育
性腺的发育使得高等动物能够产生生殖细胞,繁殖后代。
健康管理
老年人应注重健康管理,保持合理的饮食、运动和良好的生活习惯。
心理调适
老年人应学会适应身体和环境的变化,保持良好的心理状态。
社会参与
鼓励老年人积极参与社会活动,提高生活质量和社会适应能力。
老年阶段
03
CHAPTER
高等动物个体发育的影响因素
高等动物的个体发育过程受到遗传物质的控制,基因和染色体的变异会影响发育过程。
从胚胎形成到出生前的发育过程,包括各器官系统的发育和成熟。
胎儿发育
胎儿的骨骼和肌肉逐渐发育,为出生后的运动能力奠定基础。
骨骼和肌肉
胎儿的内脏器官逐渐成熟,如心脏、肺、肝脏等。
内脏器官
胎儿的神经系统逐渐发育,大脑和脊髓的结构和功能不断完善。
神经系统
胎儿发育阶段
新生儿出生后的护理和喂养,以及适应新环境的过程。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
Developmental Biology
apoptosis Self-renewal
人 约 需 70 天
减数分裂开始
精子分化(spermiogenesis)
精子的生成过程中应注意以下几点: 1、精原细胞A1是生殖干细胞,精子的发生可 持续进行。 2、分裂过程细胞质分裂不完全,形成合胞体, 细胞之间通过细胞间质桥沟通信息和物质的交 流,因而使每群细胞同步成熟。 3、精子体小,有鞭毛的蝌蚪状,具有了运动 能力和与卵子相互作用的能力。有顶体(高尔 基复合体形成的囊状结构)、鞭毛(中心粒微 管等构成),线粒体(环绕鞭毛基部),头 (细胞核染色质凝集)。大部分细胞质抛弃。
第一节 生殖细胞的起源与分化
• 一、生殖细胞在胚胎发育中的分化类型
• 1、后期分化:生殖细胞在胚胎发育后期从体细胞 分化而来(海绵、腔肠动物、扁虫等动物)。 • 2、早期分化:生殖细胞从受精卵第一次卵裂或胚 胎发育早期分出其卵裂球中含有生殖质,或由特殊 细胞形态的卵裂球分化出来(许多昆虫、棘皮动物 和脊椎动物等)。 • 在胚胎发育初期生殖细胞就已经决定的动物,生殖 细胞的前体称原生殖细胞(Primordial germ cells PGCs)。原生殖细胞(迁移)→生殖腺原基 →生殖嵴(分化)→生殖细胞。
二、生殖质与生殖细胞分化
• 原生殖细胞的起源可追溯到亲代动物的卵 细胞。有些动物的卵母细胞中已可识别决 定生殖细胞分化的生殖质(germ plasm)。 生殖质是一种具有一定形态结构的特殊细 胞质,主要由蛋白质和RNA构成,它决定着 生殖细胞的分化。
• 目前明确了线虫、果蝇、爪蟾生殖细胞分化与生 殖质的关系:
精原细胞A1
(更新的干细胞)精原细胞A2 精原细胞A1(储存的干细胞) 分裂↓ 精原细胞A3 A1 A2 分裂↓ 精原细胞A4 分裂↓ 过渡精原细胞(人类无此型,只有A、B型) 分裂↓ 精原细胞B 分裂↓ 初级精母细胞(染色体复制) ↓第一次减数分裂(同源染色体分开) 次级精母细胞 ↓第二次成熟分裂(姐妹染色体分开) 精子细胞 ↓精子的形成(变态) 精子(图P18、图与描述的不同)
• 2、决定减数分裂细胞分化方向的是性别决定基因, 如tra-I的纯合子只能发育成为产生精子的雄性个 体,而fem-I的纯合子只能发育成为产生卵子的雌 性个体。 在正常情下,秀丽新小杆线虫的生殖细胞 既能形成卵子也能形成精子。
第二节 精子的发生
• 一、精子的发生 • 二、精子发生中基因表达的调控
三、原生殖细胞的迁移
• 不同的动物原生殖细胞迁移的方式和迁移 的途径不同:
• 1、两栖类原生殖细胞的迁移
无尾两栖类原肠胚期形成的原生殖细胞(PGCs)位于 原肠腔底部内胚层中。然后,PGCs通过肠壁内胚层的侧面 迁移并进入背侧肠系膜。背侧肠系膜是连接肠壁与胚层器 官形成区域的纽带。PGCs沿背侧肠系膜往上迁移到生殖嵴, 之后PGCs与生殖嵴细胞共同形成生殖腺。 原生殖细胞定向迁移靠生殖嵴细胞内的一些迁移因子, 或是原生殖细胞主动的伸伪足变形运动。
• 鸟类与爬行类动物的原生殖细胞来源于外胚层,迁移到生 殖新月。原生殖细胞在生殖新月区内增殖,并通过血液循 环迁移到达发育中的生殖腺。 迁移机制可能是吸引原生殖细胞的化学物质。
四、减数分裂
• 原生殖细胞(有丝分裂)→生殖干细胞(减数分裂)→雌性 或雄性配子(单倍体的染色体)。 • 注意几点: 1、减数分裂前,即生殖细胞于最后一次有丝分裂之后,核 DNA继续合成并复制一次,核内DNA的含量是正常体细胞的2倍, 每条染色体内有2条中心粒相连的姐妹染色单体。 • 2、第一次减数分裂,同源染色体分配到两个子细胞中,每个 子细胞含有由两条姐妹染色单体组成的一条同源染色体。 3、第二次减数分裂则把每条染色体中的两条姐妹染色单体分 别分配到两个子细胞中去。 • 4、在减数分裂中,通过联会配对,同源染色体间遗传信息的 交换和重组,每个单倍体细胞都含有不同的染色体组合。所 以由一个个体可产生大量不同的生殖细胞。
五、生殖细胞定向分化的决定:
• 原生殖细胞具备两种发育潜能,由生殖腺内的微 环境决定发育成精子和卵子。如将雄性蝾螈的成 熟睾丸移植入雌性体内,再把蝾螈的卵巢组织转 移到睾丸,发现生殖细胞停止进行卵子发生并且 开始向精子方向分化。 • 第一种决定是生殖干细胞进入减数分裂进行配子 发生还是继续进行有丝分裂成为生殖干细胞; 第二种决定是进行减数分裂的细胞是发育成卵子 还是精子。
用雌雄同体的秀丽新小杆线虫作为实验材料证实:
• 1、决定生殖干细胞进行有丝分裂还是减数分裂的结构 是生殖腺未端的一个不分裂的细胞(称远端细胞)。靠 近该细胞的生殖干细胞进行有丝分裂,远端的侧进行减 数分裂。若激光束破坏该细胞,所有生殖干细胞均进入 减数分裂,认为远端细胞可能分泌一种能维持细胞有丝 分裂,抑制减数分裂的一种物质。
第一篇 动物个体发育模式
第一章 生殖细胞发生 第二章 受精的机制 第三章 卵裂 第四章 原肠作用:胚胎细胞的重新组合 第五章 脊椎动物的早期发育
第一章 动物生殖细胞发生
• 有性繁殖的动物个体由两部分细胞组成, 即各类体细胞和生殖细胞,二者均由受精 卵发育分化而来。
• 第一节 生殖细胞的起源与分化 • 第二节 精子发生 • 第三节 卵子发生
• 1、线虫:未受精卵 (P颗粒均匀分布于卵 质), 受精卵(P颗粒集中预定的部位), 第一次卵裂(AB大细胞和含P颗粒的P1小细 胞),第二次、第三次、第四次卵裂,P1细 胞每次分裂产生1个体细胞前体和依次产生 P1 P2 P3生殖干细胞。 P4细胞质内含有所有P颗粒,P颗粒对 于生殖细胞的分化起重要作用。(图P12)
Developmental Biology
2、线虫原生殖细胞的命运决定于Pie-1
Pie-1编码核蛋白,其缺 失导致P1-P4也向体细 胞命运分化。其作用可 能是抑制生殖细胞中基 因转录活性。
• 2、果蝇:果蝇原生殖细胞最早发现于受精 后大约90min,这些原生殖细胞位于果蝇胚 胎后端,含有特殊的极质颗粒。如果用紫 外线照射受精卵破坏集中分布于受精卵后 端胞质中的生殖质的活性,在以后的发育 中将没有生殖细胞发生。用移植生殖质的 方法,可使受体细胞分化成原生殖细胞。 • 3、爪蟾:爪蟾受精卵中也有生殖质存在, 但还没有明确哺乳动物生殖细胞分化与生 殖质的关系。
• 2、哺乳动物生殖细胞的迁移
• 哺乳动物的原生殖细胞最早见于原条后部的胚外中胚层, 先在尿囊与后肠交界处附近聚集,以后迁移到卵黄囊附 近再分成两群,沿卵黄囊的尾部通过新形成的后肠,然 后沿背侧肠系膜向上迁移,分别进入左、右两侧生殖嵴。 原生殖细胞作变形运动在两层细胞之间迁移。
• 3、鸟类和爬行类原生殖细胞的迁移
一、精子的发生
• (哺乳动物精子的发生过程) 原生殖细胞(PGC s)到达雄性胚胎的生殖腺 原基立即进入性索,直到性成熟期,性索发育成 为生精小管,其管壁是特殊的生殖上皮。 • 1、支持细胞:由性索的细胞分化成支持细胞,对 精子细胞的发育起支持、营养和保护作用。 2、生精细胞:是原生殖细胞分化而来。由基底面 →腔面依次有:
人的精子
二、精子发生中基因表达的调控
• 现已知道果蝇Y染色体的转录是控制精子发生所必需的。 果蝇的Y染色体上有5个向外伸出的DNA环,如果这5个环中 有任何一个缺失,精子尾部的结构都不正常。即使在构成 精子的所有元件都已具备的情况下也不能正确装配。显然, 在减数分裂前期转录的Y染色体特异性RNA是以后精子形成 过程所必需的。在精子发生过程中被特异性转录的基因产 物,通常是精子运动和精子与卵子结合时都必需的蛋白质。