第九章 突变、修复与重组
高中生物DNA修复与突变知识点总结

高中生物DNA修复与突变知识点总结DNA修复与突变是生物学中重要的概念,对于高中生物学知识的理解至关重要。
本文将通过总结DNA修复与突变的知识点,帮助读者更好地理解此概念。
一、DNA修复的概念与意义DNA修复是一种生物体对DNA中产生的损伤进行修复的过程。
DNA损伤的原因包括外源性因素(如辐射、化学物质)和内源性因素(如代谢产物)。
DNA修复的意义在于保持遗传物质的完整性和稳定性,确保生物正常生长、发育和繁殖。
二、DNA损伤与修复的分类1. 物理性损伤:来自于辐射,如紫外线。
2. 化学性损伤:如环境污染物、化学试剂等。
3. 内源性损伤:细胞内代谢过程中产生的氧自由基等。
4. DNA修复机制:分为直接修复、错配修复、切除修复和重组修复等。
三、DNA损伤与修复的主要类型及机制1. 基因突变:DNA碱基发生改变,包括点突变、插入突变和缺失突变等。
DNA聚合酶的错误复制和DNA损伤引起的突变是最常见的突变机制。
2. 染色体畸变:染色体结构的改变,包括染色体缺失、染色体重排等。
染色体的非同源重组和染色体断裂产生的重排引起染色体畸变。
3. 光反应和化学损伤:紫外线、化学物质等引起DNA光反应和化学反应,导致碱基损伤、碱基交联和DNA链断裂等。
4. 不同的修复机制:直接修复修复DNA中的碱基损伤,错配修复修复DNA复制过程中出现的碱基配对错误,切除修复修复DNA链断裂,重组修复修复DNA双链断裂。
四、重要的DNA修复机制1. 直接损伤修复:包括光修复和酶修复。
光修复是利用光酶修复DNA中的嘌呤二聚体损伤,酶修复则是利用特定的酶修复DNA中的碱基损伤。
2. 错配修复:包括错配识别、错配切除和错配修复等步骤,通过DNA聚合酶和联合酶的作用修复DNA中的错配碱基。
3. 切除修复:包括碱基切除修复、核苷酸切除修复和修复性DNA 重组等不同类型。
切除修复是通过核酸内切酶、外切酶等酶的作用将损坏部分切除,然后通过DNA聚合酶和联合酶的作用进行修复。
突变和修复PPT课件
第三章 突变和修复
Tay-sachs 家族性白痴病(Tay-sachs disease):是因为在其细胞 内缺乏氨基己糖脂酶,不能将神经节苷脂GM2 加工成 为GM3,结果大量的GM2累积在神经细胞中,导致中 枢神经系统退化
脱氨氧化
CMe
T
C
U
13
第二节 突变剂(续)
C→U的错误可由尿嘧啶糖 基酶系统修复
CMe→T的错误,修复系统 的修复效率较低。如果在 新一轮复制前未能完成修 复,则产生突变
14
第三节 DNA的修复
一、复制修复: 1. 尿嘧啶-糖基酶系统: 修复对象:掺入到DNA链的U(能与A配对,PolIII
无法修复) 修复过程:
27
4.SOS修复(续)
SOS反应由Rec A蛋白和Lex A蛋白相互作用引起。
Rec A被(相对分子质量为22700)称为辅蛋白酶(coprotease), 在有单链DNA和ATP存在时,Rec A蛋白被激活而促进LexA自身 的蛋白水解酶活性。
LexA蛋白是许多基因的阻遏物,当它被RecA激活自身的蛋白水 解酶活性后自我分解,使一系列SOS基因得以表达,其中包括:
6
二、突变的分类(续)
自发突变(spontaneous mutatiow) 突变率 是指在单位时间内某种突变发生的概率,即每 代每对核苷的突变概率数或每代每个基因的突 变概率。 突变频率是指在一个细胞群体或个体中,某种突变发 生的数目,即每10万个生物中发生突变体的 数目,或每百万个配子中突变的数目
第九章遗传学习题答案

第九章突变和重组机理一、名词解释1.碱基替换:一个碱基对被另一个碱基对代替。
又分2种:转换和颠换。
2.染色单体转变:减数分裂的4个产物中,有一个产物发生基因转变。
3.显性致死:只有一个致死基因就引起致死效应的,杂合态即有致死效应。
4.Muller-5品系的果蝇:X染色体上有显性棒眼基因B和隐性杏色眼基因Wa,还有一些倒位,可以抑制 Muller-5的X染色体与野生型X染色体的重组。
5、基因突变(点突变):指一个基因变成了它的等位基因。
6、自发突变:由外界环境条件自然作用或生物体内的生理生化变化发生的突变。
7、诱发突变:在特设的诱变因素(物理、化学、生物等)诱发下发生的突变。
8、显性致死:杂合态即有致死效应。
9、隐性致死:纯合态时才有致死效应,常见条件致死突变:在某些条件下是成活的,而在另一些条件下是致死的突变。
11、突变率:指生物体(微生物指每一个细胞)在每一世代中,每一基因组或每一细胞发生突变的概率。
12、易变基因:比一般基因容易突变的基因。
13、诱变剂:凡能增加突变率的物质都叫诱变剂。
14、转换:DNA分子中的嘌呤被嘌呤或嘧啶被嘧啶替换。
15、颠换:DNA分子中的嘌呤被嘧啶或嘧啶被嘌呤替换。
16、DNA修饰物:通过化学变化改造DNA分子结构的物质。
其作用与DNA复制无关。
17、染色单体转变:减数分裂的4个产物中,有一个产物发生基因转变。
18、半染色单体转变:减数分裂的4个产物中,有1个或2个产物的一半出现基因转变。
影响DNA 的一条链,分离一定发生在减数分裂后的有丝分裂中,所以叫做减数后分离。
二、选择填空1、紫外线照射主要引起( B )。
A DNA分子的断裂B 胸腺嘧啶二聚体C 尿嘧啶二聚体D DNA分子交连2、辐射剂量增加一倍,突变频率(D )。
A 不变 B增加2倍 C 增加3倍 D 增加1倍3、吖啶类分子扁平、能插入DNA相邻碱基对间,使碱基分开,从而使DNA分子双链歪斜,导致交换时出现(A )。
基因突变
突变的特点:
1. 随机性 2. 稀有性 3. 可回复性:真回复突变和第二点回复 突变
正向突变
his+
回复突变
his-
4. 多方向性 5. 通常为有害性
2007年
57.已知某一腺嘌呤缺陷型的粗糙脉胞菌受到离原 突变25个图距单位的一个抑制基因的作用后回复 为原养型,假如将这一菌株与野生型杂交,那么 后代中腺嘌吟缺陷型所占的百分数为: A.0 B.12.5% C.25% D.50% 解: ade-su- X ade+su+ 亲本型后代:ade-su-(原养型), ade+su+(原养型) 重组型后代:ade-su+(腺嘌呤缺陷型), ade+su-(原 养型), 重组型后代合计应占25%,各占12.5%。(B)
点突变的类型(以 Tyr 的密码子为例) 无义突变 DNA TAC→TAA , TAG ↓ ↓ ↓ ↓ RNA UAC UAA UAG ↓ ↓ ↓ ↓ aa Tyr Och Amb 同义突变 TAC → TAT ↓ ↓ UAC UAU ↓ ↓ Tyr Tyr 错义突变 TAC→ TCC ↓ ↓ UAC UCC ↓ ↓ Tyr Ser
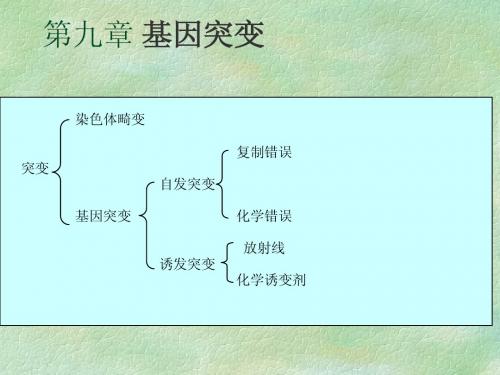
第九章 基因突变
染色体畸变 复制错误 突变 自发突变 基因突变 诱发突变 化学诱变剂 化学错误 放射线
直接修复
修复
切除修复
复制后修复
DNA Pol 3'→5'外切酶活性 光复活酶 一般切除修复——UvrABC 系统 AP 内切酶 特殊切除修复 糖基酶 GO 系统-MutM,MutY,MutT 错配修复-Dam, MutL,MutH,UvrD 重组修复-RecA SOS 修复-RecA,LexA,UvrAB,UmuC,HimA
遗传学期末总结详解演示文稿

第十三页,共59页。
第十一章 免疫遗传
1. 抗原(antigen): 凡是能够刺激机体的免疫系统产生抗体,并且能够和相应的抗体发 生特异性结合反应的物质
2. 新生儿溶血症 • 由胎儿与母体红细胞抗原不相容所引起的。 • 胎盘渗血、胎盘剥离时胎儿红细胞进入母体,母体产生全抗体IgG。IgG抗体通
2. 如何区别核外遗传和母性影响;
3. 植物的雄性不育概念及类型
4. 不育系、保持系、恢复系的关系
第二十五页,共59页。
第十三章 核外遗传
1. 核外遗传的概念和特点
细胞质内遗传物质控制的遗传现象和遗传规律。--细胞质遗传
1. 正交和反交的遗传表现不同,F1通常只表现母本的性状;
2. 与父本连续回交母本核基因可被全部置换掉,但母本细胞质基因变化不大;
5. C值悖理:
C值和进化程度之间没有严格的对应关系
6. N值悖理: 基因数目与其生物结构的复杂性不成比例的现象
7. 基因组学:
研究基因组的组成、结构和功能的学科
基因组学按研究内容:
结构基因组学
功能基因组学
蛋白质组学
第九页,共59页。
8. 结构基因组学:
是指通过基因作图、核苷酸序列分析,确定基因的组成、进行基 因定位的学科。
首先是重链发生基因重排,随后是轻链重排。
重链:胚系基因 → D-J连接 → V-DJ连接
7. 等位排斥与同种型排斥
• 等位排斥(allelic exclusion):一条染色体上Ig重链基因的有效重排,抑
制另一条同源染色体重链基因重排。 • 同种型排斥(isotype exclusion):是指两种轻链之间的排斥,轻链有κ链和λ链
基础生化-2009-第九章DNA合成

的两个子代DNA。细菌的旋转酶是Ⅱ型的代表。
(三) 引物酶(primrase)
合成RNA引物,又叫引物合成酶、引发酶。
它以单链DNA为模板,以ATP、GTP、CTP、UTP为原料,
从5→3方向合成出RNA片段,即引物。
(四) 切除引物的酶
原核细胞是DNA聚合酶Ⅰ或RNaseH(水解与DNA杂交的 RNA链)。 真核细胞是RNaseHⅠ
爬行模式
四、复制的机制
(一)θ复制(大肠杆菌为代表) (二)滚环复制 (三)D-环复制 (四)真核细胞的复制
滚环复制(rolling circle replication)是一些简单 低等生物或染色体以外的DNA复制的特殊形式。
3'
3'' 3 5'
5'
5'
五、复制的过程(大肠杆菌为例)
(一) 复制的起始第九章 DNA的生物合成• 第一节 DNA的复制
• 第二节 DNA的损伤修复
• 第三节 反转录
第一节 DNA的复制 一、复制的一般特点
1.模板、前体、Mg2+。 2.DNA的解链 3.半保留复制 4.需要引物
5.链延伸的方向
6.固定的起点(复制起始区有共有特征)
7.复制的方向(双、单)
8.半不连续复制
光修复
UV
光修复酶 (photolyase)
细菌、真菌、植物、很多脊椎动物有该酶,但胎盘类哺乳动 物没有。
2.烷基化碱基的直接修复(P601了解) 烷基转移酶,6-甲基鸟嘌呤甲基转移酶最常见。该酶以“自杀” 方式催化反应。 其活性中心的Cys残基作为甲基受体,但得到甲基就失活了。随
后被选择性降解。
二、 复制的方式——半保留复制
第九章 DNA的生物合成

主要内容
• 概述 • DNA的生物合成 • DNA的损伤与修复
概
述
遗传信息传递的中心法则
反映了从DNARNA蛋白质的遗传信息主 流,揭示了生物体内遗传信息的贮存、传递和 表达的规律。
复制
DNA
转录
RNA
翻译
蛋白质
反转录
复制
RNA (病毒)
翻译
蛋白质 (病毒)
第一节
★ 定义:
DNA的复制
复制是指以亲 代DNA为模板合成 子链DNA的过程。
四、DNA的复制过程
大致分为三阶段: 复制的起始 链的延长 复制的终止
(一)复制的起始
1. DNA解成单链
由特定蛋白质识别复制起始位点(ori)解螺 旋酶、TOPO酶及单链DNA结合蛋白的共同作用下, DNA解链,解旋,形成复制叉 2. 引发体的生成 解旋酶解开双链后引物酶进入形成引发体
3. RNA引物的合成 依赖于单链模板,由引物酶催化按碱基配对 规律合成一小段RNA引物(原核细胞引物长50-100 个碱基,真核约10个碱基)。
(4)在RNA引物上合成DNA
DNA复制的精确性(高保真复制)
DNA复制必须具有高度精确性,在大肠杆菌的细 胞DNA复制中其错误率约为1/109~1/1010,即每109~ 1010个核苷酸才出现一个错误,也就是大肠杆菌染色 体DNA复制1000~10000次才出现一个核苷酸的错误。 这么高的精确性的保证主要与下列因素有关: 1、碱基的配对规律:摸板链与新生链之间的碱 基配对保证碱基配错几率约为1/104~1/105。 2、DNA聚合酶的3’→5’外切酶活性的校对功能, 使碱基的错配几率又降低100~1000倍。 3、DNA的损伤修复系统。
(1)DNA聚合酶 即依赖于DNA的DNA聚合
突变类型与修复要点

这种突变不能引起氨基酸序列的改变。
9
碱基置换造成多肽链中一个氨基酸的改变, 但该氨基酸不能影响多肽链的正常功能, 因此不改变微生物的遗传性状。
10
移码突变是在DNA分子上的密码子中添加 或丢失一个或几个碱基,其结果造成从改 变的碱基开始所有其后的密码子碱基都往 后移动,使密码子杂乱而重新编组,使多 肽链上的氨基酸序列发生很大的改变,并 出现明显的遗传性状变异。
错义突变通常在第一或第二个碱基发生变 化时,容易引起氨基酸种类改变。
决定某氨基酸的密码子被终止密码子 (UAG)代替,因而mRNA翻译多肽链 过程中出现句号,使翻译工作中途停止, 难以完成一条完整的多肽链的合成,这种 肽链是没有活性的。
8
指DNA分子上的遗传密码由于置换而成为 新的密码子,但是这种新密码子构成的氨 基酸与原有密码子所构成的氨基酸相同。
最典型的是营养缺陷型,它从野生型基因突变形成。其特点是 由于突变而失去合成某种代谢物质的能力,如氨基酸、维生素 等,当环境中缺乏这种物质它就不能生长繁殖。反之,只有给 它补充了这种物质,才能具有正常的生命活动。
还有糖类分解发酵突变株、色素形成突变株及有益代谢产物生 产能力突变株。
突变体形成的三个步骤:
缺陷型
15
形态突变型 生化突变型 条件致死突变型 致死突变型 抗性突变型
形态突变是一种可见突变,包括:
菌落——形状大小、颜色、表面结构、孢子 数量和颜色;
细胞形态——鞭毛、荚膜、菌体形状、大小、 孢子形态和大小;
细胞结构——细胞膜透性等; 噬菌体——噬菌斑形状、大小及清晰度等。
(suppressor tRNA)。 抑制tRNA不是细胞对无义突变应答的产物,而是自发
DNA的损伤修复及突变PPT课件

着色性干皮病(xeroderma pigmentosis,XP) 是一种切除修复有缺陷的遗传性疾病。
在研究其发病机制时,发现一些相关的基 因,称为 XPA、XPB、XPC等。这些基因的表达产物起辨认 和切除损伤DNA作用的。
XP病人是由于XP基因有缺陷,不能修复紫外 线照射引起的DNA损伤,因此易发生皮肤癌。
•19
➢ DNA链断裂 脱氧核糖破坏或磷酸二酯键断开而导致DNA链断裂。 一条链断裂称单链断裂(single strand broken); DNA双链在同一处或相近处断裂称为双链断裂(double strand broken )。
•20
➢交联(binding) 同一条DNA链上或两条DNA链上的碱基间以共价
烷化剂的种类很多,常见的有甲磺酸乙酯(EMS)、 亚硝基胍(NG)和芥子气等。
•27
EMS能使鸟嘌呤的 N位置上有乙基,成为7一 乙基鸟嘌呤。与胸腺嘧啶配对,故能使G-C转换成 A-T。
烷化剂的另一作用是脱嘌呤。例如烷基在鸟嘌 呤N位上活化糖苷键引起断裂,使嘌呤从DNA链上 脱掉,产生缺口。复制时,与缺口对应的位点上可 能配上任一碱基,从而引起转换或颠换;而且去嘌 呤后的DNA容易发生断裂,引起缺失或其他突变。
•46
• 管理基因( caretaker genes) : 执行DNA的损伤修复, 维持基因组的完整性。如着色性干皮病的修复基因 XPA→XPF。
• 看门基因( gatekeeper genes) : 控制细胞信号传导, 调控细胞的增殖、分化和凋亡。如p53、patched基 因和ras等。皮肤癌的发生与看门基因突变关系密 切。
移码突变: 由于插入或缺失突变引起DNA的阅读框(ORF)
发生改变,从而产生不同蛋白质的过程。
基因突变及修复课件
将待分离突变株的原 始菌株以合适的稀释 度涂布到野生型菌株 和突变株均能生长的 平板(含完全培养基), 培养形成单菌落
通过一消毒“印章”将 平板菌落分别原位移 位到含完全培养基的 平板(c)和某一营养物 质缺乏的平板(d)
培养后对照两个平板上形成的单菌落,在完全培养基的平板 (c)上生长而营养物质缺乏的平板(d)上不生长的菌落即所需分 离的突变型,挑选菌落在完全培养基上分离纯化
基因突变及修复
一、基因突变的类型及其分离 二、基因突变的分子基础 三、DNA损伤的修复
基因突变及修复
二、基因突变的分子基础
突变
自发突变
环境因素的影响,DNA复制过程的偶 然错误等而导致,一般频率较低,通 常为10-10~10-6
诱 变 某些物理、化学因素对生物体的DNA
进行直接作用,突变以较高的频率产生
2、诱发突变(induced mutation) 常 用 诱 变 剂
(1) 碱基类似物:5-溴尿嘧啶(胸腺嘧啶类似物)和2-氨基嘌呤(嘌 呤类似物)
(2) 插入染料:插入DNA分子的碱基对,使其分开,导致DNA 在复制过程中滑动→移码突变,如溴化乙锭、吖啶橙等染料
(3) 直接与DNA碱基起化学反应的诱变剂:如亚硝酸、羟胺和烷 化剂等
基本培养基+ 少量组氨酸 +少量诱变剂
原理:(his-)菌株在不含 组氨酸的培养基中不能 生长,或只有极少数的 自发回复突变子生长, 如果回复突变率因某种 化学诱变剂(或待测物) 的作用而增加,那么这 种化学药物可判断为具 有致癌性
基因突变及修复
二、基因突变的分子基础
3、诱变剂与致癌物质——Ames试验
基因突变及修复
一、基因突变的类型及其分离
第9章__蛋白质工程中的基因突变方法

3.鸟枪合成突变法 鸟枪合成突变法
用这种方法,可在基因的一个很大 用这种方法 可在基因的一个很大 区域中同时引入多种突变. 区域中同时引入多种突变 首先将这一区域的基因序列分解成 若干可互相黏结的片段,然后合成各种 若干可互相黏结的片段 然后合成各种 所需突变的寡核苷酸,将这些寡核苷酸 所需突变的寡核苷酸 将这些寡核苷酸 与目的基因双链一起退火.一次连接即 与目的基因双链一起退火 一次连接即 得到含各种突变的基因大段序列. 得到含各种突变的基因大段序列
正 链 负链
错配部位
5)将此闭环双链 将此闭环双链DNA纯化 纯化. 将此闭环双链 纯化 6)将纯化的双链闭环 将纯化的双链闭环DNA用大肠杆菌进 用大肠杆菌进 将纯化的双链闭环 行转化,复制 其复制转化产物中含有所 行转化 复制,其复制转化产物中含有所 复制 需的突变基因. 需的突变基因
正链 负链
1.经典的寡核苷酸诱导的定点突变 经典的寡核苷酸诱导的定点突变
大体步骤: 大体步骤 1) 制备含有目的基因 即欲突变的基因) 制备含有目的基因(即欲突变的基因 目的基因 即欲突变的基因 的单链DNA(简称正链). 的单链 简称正链 简称正链
目的基因(正链) 目的基因(正链) 正链
突变点
2) 合成含有所需要突变序列的寡核苷酸引物 这种 合成含有所需要突变序列的寡核苷酸引物.这种 引物中间部分是所需突变序列,与目的基因的欲 引物中间部分是所需突变序列 与目的基因的欲 突变部位错配.其两端部分是互补区 其两端部分是互补区. 突变部位错配 其两端部分是互补区
带有目的基因 寡核苷酸 的DNA
置换后的DNA 置换后的
置换出来的 片段
1. 盒突变
盒突变是这类方法的代表. 盒突变是这类方法的代表 所谓盒,就是指与目的基因的欲突 所谓盒 就是指与目的基因的欲突 变部位相应的、 变部位相应的、由化学合成的一系列 双链寡核苷酸。 双链寡核苷酸。这一系列双链寡核苷 酸可能带有各种可能选用的突变系列。 酸可能带有各种可能选用的突变系列。 当进行突变处理时, 当进行突变处理时,根据实际需要把 各种突变盒插入目的基因中, 各种突变盒插入目的基因中,就象把 盒式录音带插入录音机一样。 盒式录音带插入录音机一样。
遗传学 遗传重组
IS
IS
IS
transposition
IS L
IS
R臂
中心区
臂
♣ 两侧的IS既可 以是IR,又可以 是DR状态 (IR多) ♣ 当两个IS组件相同 时,其中任一个都可 行使转座功能
♣ 不同时,主 要依靠一个
C、转座噬菌体 mutator phage, Mu (巨型转座子 )
以E.coli为寄主的温和型噬菌体(溶源、裂解)
玉米转座因子对胚乳颜色的影响 Ac、Ds的转座属于非复制机制
2)果蝇基因组中的转座子
果蝇的P因子有两种类型: 一类是全长P因子,长2907bp,两端有33bp的反向重复序
列(IR),有4个外显子(4个ORF),编码转座酶; 另一类为缺失型P因子,它不能编码转座酶,它的转座
依赖于全长P因子。缺失型P因子都是由活性P因子的中段 缺失衍生而来的,长度从500bp到1400bp不等。
③ 在新的位置上,转座因子两侧出现5~12bp 正向 重复序列
④ 转座过程中出现共联体(cointegrate),即两个 复制子通过共价键连接形成的一个复制子
2)非复制型转座(nonreplicative transposition) 转座子从供体一个位点转移到受体新位点处,供
体位点留下缺口,受到损伤(严重时致死)或宿主 修复系统识别修复。
Ds因子长0.4-4kb,它的中间(在转座酶基因中)有许多种长度不等的 缺失, 如Ds9缺失194bp,而Ds6则缺失2.5kb,Ds的两端也都有11bp的 反向重复序列。
Ac和Ds的末端反向重复几乎是一样的,只有一个不同之处:Ac两 端最外边的核苷酸是彼此不互补的T:G,而Ds是互补的T:A(图)。
Ac-Ds转座元件结构示意图。右边示Ac及Ds元件的单链DNA末端反向重复 配对所形成的茎环结构,这种结构可能对转座有意义
第九章微生物的遗传与变异

❖①加S菌DNA
长出S菌
活R菌
❖②加S菌DNA及DNA酶以 外的酶
❖③加S菌的DNA和DNA酶 ❖④加S菌的RNA
只有R菌
❖⑤加S菌的蛋白质
❖⑥加S菌的荚膜多糖
只有S型细菌的DNA才能将S. Pneumoniae的R型转
化为S型。且DNA纯度越高,转化效率也越高。说
明S型菌株转移给R型菌株的,是遗传因子。
(二)噬菌体感染实验
A. D. Hershey和M. Chase, 1952年
(1)含32P-DNA的一组:放射性85%在沉淀中 (2)含35S-蛋白质的一组:放射性75%在上清液中 所以,进入细胞的是噬菌体的核酸而不是蛋白质。
(三)植物病毒的重建实验
❖为了证明核酸是遗传物质,H. FraenkelConrat(1956)用含RNA的烟草花叶病毒 (TMV)进行了著名的植物病毒重建实验。
插入突变 产生染色体畸变(复制性转座子) 基因的移动和重排
第四节 基因突变及修复
一、基因突变
基因突变(gene mutation)简称突变,是 变异的一种,指生物体内遗传物质的分子 结构或数量突然发生的可遗传的变化。突 变率常在10-8~10-9范围内。
成分是蛋白质。
❖ DNA是遗传变异的物质基础的证明:1944年以后,利
用微生物为实验对象进行的三个著名实验(肺炎球菌的转化 试验、噬菌体感染试验、病毒的拆开与重建试验)
一、三个经典实验
❖ (一)经典转化实验(transformation):F.Griffith, ❖研究对象:Streptococcus pneumoniae(肺炎双球菌) ❖ S型菌株:有致病性,菌落表面光滑,有荚膜 ❖ R型菌株:无致病性,菌落表面粗糙,无荚膜
第九章 基因突变与疾病
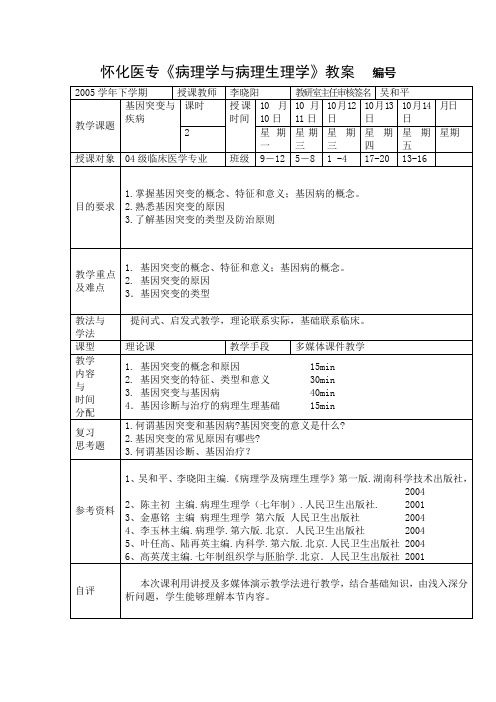
怀化医专《病理学与病理生理学》教案编号第九章基因突变与疾病(gene mutation & disease)第一节基因突变的概念和原因一、概念1.基因及其特征基因(gene)即DNA分子上一段具有遗传功能的核苷酸序列。
基因的特征:①基因能自我复制;②基因决定性状;③基因能发生突变。
2.基因突变(gene mutation) DNA分子上核苷酸序列或数目发生改变。
3.点突变(point mutation) 由一个或一对碱基发生改变引起核苷酸序列改变所致的突变。
4.缺失性和插入性突变(deletional and insertionar mutation) 即核苷酸数目改变的基因突变。
5.突变基因(multant gene) 基因突变后在原有位置上出现的新基因。
基因突变的后果生殖细胞基因突变受精卵将突变的遗传信息传给下一代(代代相传) ,即遗传性疾病。
体细胞基因突变局部形成突变细胞群(肿瘤)。
二、原因1.自发性损伤可能与DNA复制过程中碱基配对出现误差有关(自发突变)。
2.诱变剂的作用诱变剂(mutagen)是外源诱发突变的因素,种类繁多,主要有以下几种:(1)物理因素:紫外线、电离辐射等。
(2)化学因素:①烷化剂②碱基类似物: 如5-BU AT 5-BUA 5-BU A AT 5-BU 5-BU(烯醇式)(酮式)G GC碱基类似物5-BU引起DNA碱基改变示意图③其他化学诱变剂: 羟胺、亚硝酸盐等常见化学诱变剂起基因突变的机制诱变剂作用机制DNA分子改变C G-CH3 G-C T-A烷化剂C-G CH3碱基类似物(5-BU)G 5-BU A-T G-COH) C A C-G A-T羟胺类(NH2亚硝酸盐(NO) C U G-C A-T23.生物因素如:病毒、真菌、细菌等第二节基因突变的特征、类型和意义一、基因突变的特征1.多向性2.有害性3.重复性4.随机性二、基因突变的类型1.根据发生的原因分自发突变诱发突变2.根据突变的细胞不同分生殖细胞突变体细胞突变3.根据碱基改变不同分:(1) 碱基置换突变1) 同义突变:指碱基置换后,密码子虽发生改变,但其编码的氨基酸并未改变,并不影响蛋白质的功能,不发生表型的变化,即改变前后的密码子为同义密码。
《基因突变和基因重组》教案

《基因突变和基因重组》教案第一章:基因突变概述1.1 基因突变的定义1.2 基因突变的原因1.3 基因突变的特点1.4 基因突变的意义第二章:基因突变的发生机制2.1 点突变2.2 插入突变2.3 缺失突变2.4 倒置突变2.5 基因重排第三章:基因突变与疾病3.1 遗传性疾病3.2 肿瘤3.3 先天性疾病3.4 基因突变与个体差异第四章:基因重组概述4.1 基因重组的定义4.2 基因重组的类型4.3 基因重组的特点4.4 基因重组的意义第五章:基因重组的发生机制5.1 同源重组5.2 非同源重组5.3 转座重组5.4 病毒介导的重组5.5 人工基因重组技术第六章:基因突变与生物进化6.1 基因突变在生物进化中的作用6.2 突变-选择平衡理论6.3 基因流与基因漂变6.4 现代生物进化理论视角下的基因突变第七章:基因重组的应用7.1 基因工程7.2 基因编辑技术(如CRISPR-Cas9)7.3 基因治疗7.4 基因重组在农业中的应用7.5 基因重组在医学研究中的应用第八章:基因突变与法律伦理问题8.1 基因隐私权8.2 基因歧视8.3 基因治疗的伦理问题8.4 基因编辑技术的伦理争议8.5 相关法律和国际协议第九章:基因突变与基因重组的研究方法9.1 分子生物学技术9.2 遗传学实验方法9.3 基因组学分析9.4 生物信息学在基因研究中的应用9.5 实验设计与数据分析第十章:综合练习与案例分析10.1 基因突变和基因重组的概念辨析10.2 案例分析:遗传疾病的基因诊断与治疗10.3 问题解决:设计一个基因编辑实验10.4 小组讨论:基因技术在未来的应用前景10.5 复习测验:基因突变和基因重组的知识点回顾教案的内容应该根据学生的学习水平和课程的具体要求进行调整,确保教学内容的深度和广度适合学生的情况。
教案中应该包含互动环节和实践活动,以提高学生的参与度和理解力。
重点和难点解析重点环节1:基因突变的特点和意义基因突变的特点包括随机性、低频性、多数有害性等。
第九章 突变、修复与重组

-5个核苷酸,5’切割点距损伤位点8个核苷酸。
UvrD与5’端的切点结合。UvrD是一种解旋酶(又称DNA
helicase II),它解开两个切点之间的DNA双螺旋,
导致一段短的带有损伤的ssDNA和UvrC被释放出来,
此时UvrB仍结合于另一条单链DNA分子上,可能是防
止单链被降解,也可能是指导DNA聚合酶I与缺口的3’ OH结合,合成一段新的核苷酸片断填补缺口,同时释
一条子代DNA分子上的C ∶ G被 T∶ A所取代。细胞在 进化过程中,形成了多种修复机制,以保证在损伤阻 遏复制或者产生突变之前就识别并修复损伤。
一、光 复 活( Photoreactivation )
在可见光存在的情况下,DNA光解酶(DNA photolyase) 把环丁烷嘧啶二聚体分解为单体。DNA光解酶,又称光 复活酶(photoreactivating enzyme)。光解酶在暗 中结合到环丁烷二聚体上,吸收300~350 nm的光后被 激活,裂解二聚体后与DNA分子脱离。从光复活修复过 程可以看出,光解酶不是将嘧啶二聚体替换掉,而是 将两个嘧啶环之间的非正常化学键切开,恢复到原来 的形式。由于这种修复作用只在可见光下才可发生, 所以这种修复机制称为光复活。
放出UvrB,最后一个磷酸二酯键由DNA连接酶催化形
成。
核苷酸的切除修复
DNA解螺旋酶
核苷酸切除修复(2)
uvrB、uvrA 蛋白结合 uvrD 切除
碱基替换:
根据碱基变化的不同,可以分为:
转换(transition):嘌呤与嘌呤之间,或嘧啶 与嘧啶之间的替换 颠换(transversion):嘌呤与嘧啶之间的替换。
缺失、插入 缺 失 和 插 入 常 导 致 移 码 突 变 ( frameshift mutations),结果产生截短的或异常的多肽链。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
A· TG· transition C
2. 脱氨剂 (Deamination agents)
除去碱基上的氨基,改变其配对性质
3. 烷化剂 (Alkylating )
使DNA中的碱基发生烷化作用,引起特异性错配,或脱嘌呤。 包括芥子气,甲磺酸已酯(EMS),亚硝基胍(NG)等。如EMS,
自发损伤(spontaneous lesions)
自然产生的DNA损伤引起突变
1. 脱嘌呤:最为常见,DNA分子中碱基和脱氧核糖间的糖 苷键受到破坏,从而引起鸟嘌呤(G)或腺嘌呤(A)从DNA分子 上脱落下来。
常引起缺失或插入突变
2. 氧化性损伤:个体自然产生的氧化基,氧化物如超氧 自由基,氢氧自由基及过氧化基等,能对DNA造成氧化性损 伤,引起突变。
填补上。它可以修复一系列损伤,包括环丁烷二聚
体、6-4光产物和链间交联等引起的DNA变形 (major distortion)。尽管这些损伤也可以通过 途径进行修复,但NER是它们的主要修复手段。其 他能够引起DNA产生显著变形的损伤也可以通过该
途径进行修复。但是,这种机制不能修复DNA上的
错配碱基,以及仅造成DNA微小变形的碱性类似物 和甲基化碱基。
-5个核苷酸,5’切割点距损伤位点8个核苷酸。
UvrD与5’端的切点结合。UvrD是一种解旋酶(又称DNA
helicase II),它解开两个切点之间的DNA双螺旋,
导致一段短的带有损伤的ssDNA和UvrC被释放出来,
此时UvrB仍结合于另一条单链DNA分子上,可能是防
止单链被降解,也可能是指导DNA聚合酶I与缺口的3’ OH结合,合成一段新的核苷酸片断填补缺口,同时释
放出UvrB,最后一个磷酸二酯键由DNA连接酶催化形
成。
核苷酸的切除修复
DNA解螺旋酶
核苷酸切除修复(2)
uvrB、uvrA 蛋白结合 uvrD 切除
•
非条件型突变: 在任何条件下均可表现的突变 条件型突变: 突变的表现 = 突变基因型 + 诱导条件
根据突变的方式:
碱基替换(base substitution),DNA分子中一个碱 基被另一个碱基替代 又称点突变 插入(insertion),DNA序列中插入一个或多个碱基 缺失(deletion),DNA序列上缺失一个或多个碱基 重复(duplication),一段碱基序列发生一次重复 易位(translocation),一段碱基序列从原来的位 置移出,并插入到基因组的另一位置 倒位(inversion),一段碱基序列发生倒转,但仍 保留在原来的位置上
二、烷基的转移
一些酶可将烷基从核苷酸转移到自身的多肽链上,
例如,在人类细胞中发现有一种O6-甲基鸟嘌呤甲基
转移酶,能直接将DNA链鸟嘌呤O 6 位上的甲基移到 蛋白质的半胱氨酸残基上而修复损伤的DNA。
三、核苷酸切除修复
核苷酸切除修复需要移去一段包括损伤在内的核 苷酸序列,然后再通过DNA合成把余下的单链缺口
4 通读(reading through):碱基替换使终止密码子突
变为编码氨基酸的密码子,转录出的mRNA在翻译时不能适时 终止。
按突变发生的原因分类 • 自发突变(spontaneous mutation):在自然状况 下发生的突变。
• 诱发突变(induced mutation):外界因素诱发 引起的突变。
改变了野生型性状的突变,使性状由 野生型变为突变型。
回复突变(reverse mutation):
使突变型性状恢复到野生型性状的 突变。 大多数回复突变都发生在另一位点。因此,这样的突变 并未恢复野生型的序列,只是其表型被抑制了。第二点 突变可以发生在同一基因上,也可以发生在不同的基因 上,前者称为基因内抑制,后者称为基因间抑制。
使G的N位置带有已基(已烷化),成为7-已基鸟嘌呤,不与C配
对而与T配对,使G-C转换成A-T。
4.嵌入剂(intercalating agents) 吖啶橙(acridine orange)、原黄素( proflavine) 和溴化乙啶(ethidium bromide)
吖啶类化合物是一种平面三环分子, 其大小和形状与一个碱基对相似, 插入DNA分子中两个相邻的碱基之 间,使得原来相邻的碱基对分开一 定的距离,致使DNA在复制时增加
1. 基因内抑制
氨基酸上所带的电荷影响蛋白质构象
氨基酸侧链的大小影响构象
2.基因间抑制
如:无义突变的表型 可被tRNA基因的突变 所抑制
基因间抑制常发生在 tRNA或tRNA功能相 关基因上。
错义抑制突变( Missense suppressor )
GGA CCU
Gly
AGA UCU
Arg Gly
碱基替换:
根据碱基变化的不同,可以分为:
转换(transition):嘌呤与嘌呤之间,或嘧啶 与嘧啶之间的替换 颠换(transversion):嘌呤与嘧啶之间的替换。
缺失、插入 缺 失 和 插 入 常 导 致 移 码 突 变 ( frameshift mutations),结果产生截短的或异常的多肽链。
CCU
UCU
Gly
Gly
基因间移码抑制突变
基因内和基因间的错义(无义)、移码抑制突变均 由相应的错义(无义)、移码突变抑制
突变热点(hot spot),
突变可以发生在基因组中的任一位点。但是在基因组中,也 存在一些位点,这些位点发生突变的几率比随机分布所估计的
要高出许多,可能是预期的10倍或100倍,这些位点被称为突
第九章 突变、修复与重组
第一节 突变
第二节 修复 第三节 重组
本章要点:
• 突变的概念、类型 • DNA修复的主要方式 • 了解重组的分子基础
第一节:突变(mutation) 一、突变的主要类型
突变是DNA碱基序列水平上的永久的、可遗传的改变。 广义的突变包括染色体畸变和基因突变 狭义的突变通常指基因突变 根据突变发生的细胞类型、对基因功能的影响、突变的方 式、发生的原因等可对突变进行不同的分类
变热点,发生在热点上的突变常常是相同的。 大多数突变热点是DNA分子中的5-甲基胞嘧啶位点。
5mC处的突变率明显高于其它碱基处
增变基因(mutator gene): 突变后可使整个基因组中的突变率明显上升 的基因。
增变基 因大多 与DNA 复制或 修复有 关。
第二节 DNA的修复
一系列物理或化学因素可以对DNA造成化学损伤。这
修复过程需要多种酶的一系列作用,其中包括UvrA,
UvrB, UvrC 和 UvrD。由2个UvrA亚基和1个UvrB亚基
构成的复合体,非特异性地结合在DNA分子上,并沿
DNA分子滑动,对DNA进行扫描,其中UvrA负责检测螺 旋中的扭曲,一旦抵达扭曲部位,UvrA就会退出复合 体。UvrB募集UvrC, UvrC具有核酸内切酶活性,它切 断损伤位点两侧的磷酸二酯键,3’切割点距损伤位点4
(1)物理诱变剂 • 紫外线 • X-射线和γ-射线
紫外线:波长为100~400 nm的电磁辐射,非电离辐射, 直接作用于DNA。 嘧啶二聚体 胸腺嘧啶 X-射线和γ-射线:电离辐射 • 直接作用于DNA,对DNA产生损伤 产生多 种突变,并且常常造成DNA重排,例如缺失、 倒转和移位。 • 作用于水分子以及其他分子产生离子和自由基, 自由基对DNA分子产生广泛损伤。
一条子代DNA分子上的C ∶ G被 T∶ A所取代。细胞在 进化过程中,形成了多种修复机制,以保证在损伤阻 遏复制或者产生突变之前就识别并修复损伤。
一、光 复 活( Photoreactivation )
在可见光存在的情况下,DNA光解酶(DNA photolyase) 把环丁烷嘧啶二聚体分解为单体。DNA光解酶,又称光 复活酶(photoreactivating enzyme)。光解酶在暗 中结合到环丁烷二聚体上,吸收300~350 nm的光后被 激活,裂解二聚体后与DNA分子脱离。从光复活修复过 程可以看出,光解酶不是将嘧啶二聚体替换掉,而是 将两个嘧啶环之间的非正常化学键切开,恢复到原来 的形式。由于这种修复作用只在可见光下才可发生, 所以这种修复机制称为光复活。
自发突变的分子基础
• DNA复制错误
• 自发损伤
• 转座因子
DNA复制错误
• 遗传物质是DNA,DNA复制是半保留复制,如果发生 错误,引起碱基替换(base substitution), 造成 DNA遗传信息的改变。从而导致基因突变。
正常情况下,A-T配对,氨基态的腺嘌呤(A)只与胸 腺嘧啶(T)配对,但有时可转变成稀有的亚氨基形式, 可以与胞嘧啶配对,形成A-C, 再经一次复制,DNA分 子中的A-T对变成了G-C对.
根据突变发生的细胞类型:
• 生殖细胞突变(Germline mutation) • 体细胞突变(Somatic mutation)
可遗传给后代
仅表现于当代,癌症
根据对基因功能的影响
• 功能失活突变(loss of function mutation):
使基因功能失活或显著下降的突变。 • 功能获得突变(gain of function mutation): 使基因功能增强或不受抑制的突变。 根据突变的表达类型 •
(2)化学诱变剂
1
碱基类似物:
与碱基结构类似,可替代正常碱基掺入DNA 分子,引起碱基替换。 如5-溴尿嘧啶(BU)是胸腺嘧啶T的类似物,可 掺入DNA分子中。 酮式BU与A配对,而烯醇式与G配对 容易引起G-C对与A-T对的互相转换。。
碱基类似物还有5-溴脱氧尿苷(BrdU),5-尿嘧啶,5-氯尿 嘧啶及其脱氧核苷,2-氨基嘌呤(2-AP)等
正常人 红细胞