兴奋性氨基酸受体阻断剂
兴奋性氨基酸受体拮抗剂

兴奋性氨基酸受体拮抗剂发明背景在哺乳动物的中枢神经系统(CNS)中,神经冲动的传递受传送神经元释放的神经递质与接受神经元上表面受体之间的相互作用调控,这种相互作用导致这种接受神经元兴奋。
L-谷氨酸是CNS中最丰富的神经递质,介导哺乳动物体内主要的兴奋途径,因此被称为兴奋性氨基酸(EAA)。
应答于谷氨酸的受体称为兴奋性氨基酸受体(EAA受体)。
兴奋性氨基酸受体分为两大类。
直接与神经元细胞膜中的开放性阳离子通道结合的受体称为“离子型”。
这种类型受体至少已分为三种亚型,它们根据对选择性激动剂N-甲基-D-天冬氨酸(NMDA)、α-氨基-3-羟基-5-甲基异噁唑-4-丙酸(AMPA)和红藻氨酸的去极化作用而定义。
分子生物学研究已经确认AMPA受体是由亚单位(GluR1-GluR4)组成,它们可以组装形成功能性离子通道。
已经鉴定出五中红藻氨酸受体,它们被分为高亲和性(KA1和KA2)或低亲和性(由GluR5,GluR6,和/或GluR7亚单位组成)两类。
第二大类受体是G-蛋白偶联或第二信使连接的“代谢型”兴奋性氨基酸受体。
该第二大类受体与多个第二信使系统结合,从而能够增强磷酸肌醇的水解、激活磷脂酶D、增加或降低cAMP的形成以及改变离子通道的功能。
这两类兴奋性氨基酸受体可能不仅介导正常突触沿兴奋性途径的传递,而且还参与发育和整个生命过程的突触连接的改变。
过度或不适当地刺激兴奋性氨基酸受体会以称为兴奋性毒性的机制方式导致神经元细胞损伤或损失。
已经有人提出,该过程在许多神经性疾病或病症中介导神经元变性。
这种神经元变性的医学后果对这些变性神经病变过程的缓解具有重要的治疗意义。
例如,兴奋性氨基酸受体的兴奋性毒性与多种神经性疾病的病理生理学有关,包括心脏旁路手术和移植后大脑功能性缺氧或缺血(cerebral deficit)、中风、脑缺血、创伤或炎症引起的脊髓损伤、产期缺氧、心博停止和低血糖性神经损伤的病因学。
此外,兴奋性毒性还与慢性神经变性疾病包括阿耳茨海默氏病、杭廷顿氏舞蹈病、遗传性共济失调、艾滋病诱发的痴呆、肌萎缩性侧索硬化、特发性和药物引发的帕金森病以及眼部损伤和视网膜病有关。
神经递质、受体、激动剂和拮抗剂的类型 (2)

2、神经递质、受体、激动剂和拮抗剂的类型
神经递质受体激动剂拮抗剂胆碱类:
乙酰胆碱M-受体:M1-M5
N-受体:N1、N2
M:
毒菌碱
胆碱酯酶抑制剂:
新斯的明
毒扁豆碱
腾喜龙
有机磷脂类
三乙基胆碱
4-吡啶
抑制Ach释放:
肉毒毒素
河豚毒
单胺类: (1
a
b多巴胺c肾上腺素
(2)吲哚胺: 5-羟色胺
血清紧张素5-羟色胺受体:
5-HT1—5-HT7受
体
多巴胺激动剂:
左旋多巴
苯丙胺(安非他
明)
可卡因
哌甲酯(利他灵)
α:
异丙肾上腺素、NE
β:NE、E
多巴胺拮抗剂:
AMPT
氯内嗪
氯氮平
心得安
心得平
心得静
氨基酸类:
(1)抑制性氨基酸谷氨酸门控离子
通道受体:
谷氨酸激动剂:
NMDA
AMPA
谷氨酸拮抗剂:
AP5
酒精
类:
甘氨酸
(2)兴奋性氨基酸类:
谷氨酸
天冬氨酸NMDA受体
非NMDA受体
(AMPA受体、KA
受体)
G蛋白耦联谷氨受
体:
红藻氨酸
γ-氨基丁酸激
动剂:
毒蝇蕈醇
巴氯芬
苯二氮卓类
PCP
γ-氨基丁酸拮
抗剂:
荷牡丹碱
CGP335348
印防己毒素
多肽类:阿片肽类胃肠肽类激肽类
其他:
前列腺素组胺
内皮源性舒张因子(NO、CO)
核苷类核苷类的阻断剂:咖啡因
NO的拮抗剂:
L-NAME。
兴奋性氨基酸受体拮抗剂对清醒大鼠脑缺血再灌注损伤的保护作用

s d .Fv / g r1 g k P , 2 g k r 0mgk e mie a d1 g k r 0mgk B X w r g e t y i mgk 0m / gC P 5m / go / g t n , n 5m / go / gN Q ee i n u e o 5 ka 3 v
维普资讯
实用医学 杂志 2 0 0 7年 第 2 3卷第 7期
兴奋性氨基酸受体拮抗剂对清醒大 鼠脑缺血 再灌 注损 伤 的保护作用
李 金 宝 王 晓琳 陈 辉 熊源 长 邓 小 明
摘 要 目的 : 察兴奋性 氨基酸 受体拮 抗剂对 清醒 大鼠全 脑缺血再灌 注损伤的影 响。方法 : 用我们改进 观 采 的清醒 大鼠脑缺 血模型 , 于缺 血前 1 n 再灌 注 6h分 别腹 腔注射 5 / g或 1 / g的竞争性 N 甲基一 5 、 mi k mg 0mg k 一 D天
用 . 且 这 种 保 护 作 用不 是 由 于 体 温 下 降 所 致酸 拮 抗剂 保护
Efet fe ct tr mio a i e e tr a tg n s n b an ic e c r p ru in d ma ei as L f cso x i o y a n cd r c p o n a o it o r i s h mi e e fso a g n r t I且nb o, a s —o
W NG Xa— n C E u , ING u nzag D N a— i . D p r et o eteioy te f l t A i l ,H N H iX O Y a —h , E G Xi r n oi n ou g eat n f Ans s l , A ie m h og h i ad C aga Ho il te eodMitr dcl nvr t, hnh i 04 3 C ia h hi s t h cn la Me i i sy S ag a 2 0 3 , hn n p ao f S iy aU ei
药理学名词解释人卫第七版

药理名词解释1、药理学(pharmacology):研究药物与机体相互作用及作用规律的学科,既研究药物对机体的作用及作用机制(药物效应动力学),也研究药物在机体的影响下所发生的变化及其规律(药物代谢动力学)。
2、首过消除(first pass elimination):从胃肠道吸收入门静脉系统的药物在到达全身血液循环前必先通过肝脏,如果肝脏对其代谢能力强,或由胆汁排泄的量大,则使进入全身血循环内的有效药物量明显减少,这种作用称为~。
3、再分布(redistribution):首先分布到血流量大的脑组织发挥作用,随后由于其脂溶性高又向血流量少的脂肪组织转移,以致病人迅速苏醒,这种现象称为药物在体内的~。
4、肝肠循环(enterohepatic cycle):被分泌到胆汁内的药物及其代谢产物经由胆道及胆总管进入肠腔,然后随粪便排泄出去,经胆汁排入肠腔的药物部分可再经小肠上皮细胞吸收经肝脏进入血液循环,这种肝脏、胆汁、小肠间的循环称~。
5、一级消除动力学(first-order emilination kinetics):是体内药物在单位时间内消除的药物百分率不变,也就是单位时间内消除的药物量与血浆药物浓度呈正比,也称线性动力学(linear kinetics)。
TD:大多数药物属于、比率恒定、半衰期=0.69/k。
6、零级消除动力学(zero-order emilination kinetics):是药物在体内以恒定的速率消除,即不论血浆药物浓度高低,单位时间内消除的药物量不变也称非线性动力学(nonlinear kinetics)。
TD:少数药物属于、半衰期可变。
7、稳态浓度(steady-state concentration,Css):等量等间隔连续多次给药,经4~5个半衰期后,血药浓度稳定在某一水平,称为~,亦称为坪值。
TD:消除药量与吸收药量相等、水平波动在有效浓度和中毒浓度之间、坪值波动与每次剂量和给药间隔呈正比、达坪时间与半衰期呈正比、首剂加倍立即达坪。
谷氨酸受体拮抗剂对培养大鼠海马神经元活力的影响

[文章编号]1006-2440(2012)06-0548-04谷氨酸是一种兴奋性氨基酸,主要有N-甲基-D-天冬氨酸(NMDA )和AMPA 两种亚型。
以往研究表明它对神经元具有兴奋性毒性作用[1],人们常用其受体拮抗剂来阻断这种毒性作用[2]。
但近年研究发现,谷氨酸的受体拮抗剂虽阻断了谷氨酸的毒性,但自身似乎也对细胞有一定的毒性,这种作用还不为人们所熟知[3]。
本研究分别使用NMDA 和AMPA 的拮抗剂作用于培养海马神经元,旨在探讨不同受体拮抗剂对神经元的影响。
1材料与方法1.1材料(1)动物:孕18天SD 胎鼠30只由南通大学动物实验中心提供。
(2)主要试剂:DMEM/F12培养液、Neurobasal 无血清神经培养基、B27、0.5%胰蛋白酶(Gibco 公司),台盼蓝(trypan )、四甲基偶氮唑盐(MTT )、酸化异丙醇、卓西平马来酸盐(MK-801)、艾芬地尔(ifenprodil )、CGP-37849、L-701.324、NBQX 和HA-966(上海宝曼生物公司);TUNEL 试剂盒(德国罗氏公司);多聚赖氨酸、阿糖胞苷(Sigma 公司)。
*[作者简介]李爱红,女,汉族,生于1970年7月,江苏如皋人,硕士,副主任医师。
研究方向:颅脑损伤及脑保护治疗。
通信作者:郭爱松,E-mail:guoaisong@谷氨酸受体拮抗剂对培养大鼠海马神经元活力的影响李爱红1*,郭爱松2,陈鑫1(南通大学附属医院1神经内科;2康复医学科,江苏226001)[摘要]目的:观察N-甲基-D-天冬氨酸受体拮抗剂卓西平马来酸盐、艾芬地尔、CGP-37849、L-701.324、HA-966和AMPA 受体拮抗剂NBQX 分别对培养大鼠海马神经元细胞活力的影响。
方法:取孕18天的SD 胎鼠海马神经元细胞原代培养,培养的细胞在加入上述药物后,在培养第2天和第7天行台盼蓝染色观察细胞存活情况,四甲基偶氮唑盐法检测细胞存活率,TUNEL 法检测细胞凋亡。
兴奋性氨基酸神经毒作用和神经系统疾病

兴奋性氨基酸神经毒作用和神经系统疾病兴奋性氨基酸(excitatory amino acids,EAA)是指具有2个羧基和1个氨基的酸性游离氨基酸包括谷氨酸(Glu)、天门冬氨酸(Asp),是中枢神经系统的兴奋性神经递质,尤其谷氨酸是中枢神经系统含量最高、分布最广、作用最强的兴奋性神经递质。
兴奋性氨基酸(Excitatory amino acid,EAA)是广泛存在于哺乳类动物中枢神经系统的正常兴奋性神经递质,参与突触兴奋传递,学习记忆形成以及与多种神经变性疾病有关。
缺血、缺氧、创伤、中毒等因素能触发中枢神经系统的EAA过度兴奋,在能量代谢失衡的基础上,异常堆积,产生神经毒性作用。
1957年Lucas等首次报告给小鼠注射大剂量谷氨酸可引起视网膜神经元变性,但那时没有引起足够的反响。
70年代Olney等发现给未成熟动物全身注射谷氨酸或其它兴奋性氨基酸(EAA),所有不被血脑屏障保护的脑区内神经元都发生了变性,而且不同EAA引起几乎相同的神经细胞病理形态特征,表现为突触后EAA受体所在处的树突胞体膜结构的破坏。
给成年动物局部脑区注入EAA,可引起相似的损害。
从结构和活性关系研究中观察到不同EAA及其类似物的兴奋作用和神经毒作用。
5-羟色胺还能增强记忆力,并能保护神经元免受“兴奋神经毒素”的损害。
因此充足的5-羟色胺确实能在老化过程中防止脑损害发生。
兴奋性氨基酸包括谷氨酸和天冬氨酸等。
其中谷氨酸是哺乳动物中枢神经系统中主要的兴奋性神经递质,其可对神经元的结构和功能产生长期影响,且谷氨酸介导的兴奋性信号传导可以影响哺乳动物的脑功能,包括认知,记忆和学习功能[1‐2]。
天冬氨酸与各种神经元活动密切相关,包括视觉生理学,神经形成,学习和记忆过程[5]。
在动物大脑中,天冬氨酸和谷氨酸是钙依赖性的兴奋性神经递质,且通过与谷氨酸受体结合发挥作用[4]。
谷氨酸受体可分为离子型受体和代谢型受体。
目前有3种类型的离子型谷氨酸受体,根据其原型激动剂分别命名为N‐甲基‐D‐天冬氨酸(N‐ methyl‐D‐aspartic acid ,NMDA ]受体,α‐氨基‐3‐羧基‐5‐甲基异恶唑‐4‐丙酸[2‐ami‐no‐3‐(3‐hydroxy‐5‐methyl‐isoxazol‐4‐yl ) propanoic acid ,AMPA]受体和海人酸(kainate ,KA)受体[5];代谢型谷氨酸受体(metabotropic glutamate recep‐tor ,mGLUR )分为不同的8个亚型:mGLUR1、mGLUR2、mGLUR3、mGLUR4、mGLUR5、mGLUR6、mGLUR7、mGLUR8。
药理学名词解释人卫第七版
药理名词解释1、药理学(pharmacology):研究药物与机体相互作用及作用规律的学科,既研究药物对机体的作用及作用机制(药物效应动力学),也研究药物在机体的影响下所发生的变化及其规律(药物代谢动力学)。
2、首过消除(first pass elimination):从胃肠道吸收入门静脉系统的药物在到达全身血液循环前必先通过肝脏,如果肝脏对其代谢能力强,或由胆汁排泄的量大,则使进入全身血循环内的有效药物量明显减少,这种作用称为~。
3、再分布(redistribution):首先分布到血流量大的脑组织发挥作用,随后由于其脂溶性高又向血流量少的脂肪组织转移,以致病人迅速苏醒,这种现象称为药物在体内的~。
4、肝肠循环(enterohepatic cycle):被分泌到胆汁内的药物及其代谢产物经由胆道及胆总管进入肠腔,然后随粪便排泄出去,经胆汁排入肠腔的药物部分可再经小肠上皮细胞吸收经肝脏进入血液循环,这种肝脏、胆汁、小肠间的循环称~。
5、一级消除动力学(first-order emilination kinetics):是体内药物在单位时间内消除的药物百分率不变,也就是单位时间内消除的药物量与血浆药物浓度呈正比,也称线性动力学(linear kinetics)。
TD:大多数药物属于、比率恒定、半衰期=0.69/k。
6、零级消除动力学(zero-order emilination kinetics):是药物在体内以恒定的速率消除,即不论血浆药物浓度高低,单位时间内消除的药物量不变也称非线性动力学(nonlinear kinetics)。
TD:少数药物属于、半衰期可变。
7、稳态浓度(steady-state concentration,Css):等量等间隔连续多次给药,经4~5个半衰期后,血药浓度稳定在某一水平,称为~,亦称为坪值。
TD:消除药量与吸收药量相等、水平波动在有效浓度和中毒浓度之间、坪值波动与每次剂量和给药间隔呈正比、达坪时间与半衰期呈正比、首剂加倍立即达坪。
2021年执业医师继续教育二次海默病及药物治疗答案

2021年执业医师继续教育二次海默病及药物治疗答案1、胆碱酯酶抑制剂可引起剂量依靠性(幻觉、激惹、震颤、意识模糊、瞳孔缩小、视物模糊、心动过缓以及胃肠出血等)的效应,此种效应属于是(单项选择)BA.烟碱能效应B胆碱能效应C.抗胆碱能效应D.肾上腺素能效应2、与阿尔茨海默病发病的最亲密相关和最为风险的基因是(单项选择)AA载脂蛋白E-4(ApoE4)基因B血管紧急素转换酶(ACE)基因C淀粉样蛋白前体蛋白基因D早老素-1(PS-1)3、属于兴奋性氨基酸受体阻断剂的抗痴呆药是(单项选择)CA多奈哌齐B多塞平C美金刚D阿托品4、临床提倡对阿尔茨海默病者采纳多种不同作用机制的抗痴呆药的联合治疗,获益大于单药治疗,其优势机制在于(单项选择)BA抵消各药的不良反应B显著改善患者认知功能C.节约药物治疗成本D增加患者的用药依从性5、下列不属于胆碱酯酶抑制剂的的抗痴呆药是(单项选择)DA利斯的明B加兰他敏C石杉碱甲D艾地苯醌6、阿尔茨海默病的起病年龄多是(单项选择)CA在30岁以上B在50岁以上C在60岁以上D在90岁以上7、下列不属于脑血流促进剂的抗痴呆药是(单项选择)A A多奈哌齐B艾地苯醌C尼麦角林D氟苯桂嗪8、对阿尔茨海默病患者禁用的药品类别是?(单项选择)B A抗组胺药B抗胆碱药C抗精神病药D抗抑郁药9、雌激素可以应用于阿尔茨海默病患者,所采纳其的作用机制是(单项选择)BA促进女性特征的形成B促进乙酰胆碱的生成C促进尿酸的夜间排泄D促进肾上腺素的合成10、银杏叶提取物用于对抗痴呆的作用可归结于(单项选择)DA促进大脑组织对磷脂和氨基酸的利用B与乙酰胆碱竞争性结合乙酰胆碱酯酶C阻断谷氨酸浓度病理性上升所致的神经元损伤D改善脑组织循环和代谢,增加脑血流量。
脑梗死诊疗规范

脑梗死诊疗规范脑梗死是各种原因造成的脑动脉血管堵塞,导致该血管支配区域的脑组织因缺血、缺氧而发生坏死,并产生相应的神经功能缺损的症状和体征。
这是急性脑血管病常见的一个类型,约占全部急性脑血管病的70%左右。
通常脑梗死分为动脉粥样硬化性血栓性脑梗死、脑栓塞、腔隙性脑梗死、脑分水岭梗死四种类型。
【诊断标准】(一)临床表现1.起病年龄:中、老年居多,风心病房颤为病因者年纪较轻,动脉炎性脑梗死以中青年多见。
2.起病方式:脑血栓形成多于安静状态起病,1~2天症状达到高峰;脑栓塞起病急骤,数秒钟至数分钟症状达到高峰。
3.症状:(1)颈内动脉系统脑梗死表现为急性发生的一侧肢体力弱、麻木、言语困难等,椎一基底动脉系统脑梗死主要表现为眩晕、恶心、呕吐、耳鸣、讲话不清、吞咽困难和肢体力弱等。
(2)多数患者头痛不重,但大面积脑梗死、脑水肿明显者头痛严重,可伴恶心呕吐。
4.体征:(1)神志清楚,严重者可有嗜睡到昏迷等不同程度的意识障碍。
(2)颈内动脉系统脑梗死,主要体征有偏瘫、偏身感觉障碍、偏盲,病灶在优势半球者有失语;椎一基底动脉系统脑梗死主要体征有眼球震颤、构音不清、共济失调、交叉性瘫痪。
(二)辅助检查1.头CT扫描脑梗死发生12~24小时以后,可显示低密度梗死灶,早期CT扫描未见异常可以除外脑出血,但不能除外急性脑梗死,应于起病24小时以后复查CT扫描。
2.头MRI脑梗死亚急性期,梗死部位表现为异常的长T1、长T2信号。
头MRI显示病灶时间早,一般梗死3小时后即可显示病灶。
但除非怀疑有小脑、脑干梗死,一般急性脑梗死应首选CT扫描,不需做头颅MRI。
脑干、小脑梗死头颅CT由于受到后颅凹骨骼伪影的干扰,往往不能很好显示病灶,而MRI可以清晰地显示病灶。
3.其他实验室检查应寻找及确定发生脑梗死的危险因素,如高血糖、高血脂、血液高凝状态、高粘滞状态等。
4.TCD检查可显示脑部大血管血流改变情况。
5.颈部超声可显示颈内动脉或椎动脉有无狭窄、斑块及狭窄程度。
电大药学专调剂学名词解释汇总

电大药学专调剂学名词解释汇总1. 药理学:是研究药物与机体(含病原体)相互作用及作用规律的学科。
2. 药物效应动力学:药理学即研究药物对机体的作用即作用机制,即药物效应动力学,又称药效学。
3. 药物代谢动力学:药理学也研究药物在机体影响下所发生的变化及规律,即药物代谢动力学,又称药动学。
4. 吸收:药物自用药部位进入血液循环的过程称为吸收。
药物只有经吸收后才能发挥全身作用。
5. 分布:药物一旦被吸收进入血液循环内,便可能分布到机体的各个部位和组织。
药物吸收后从血循环到达机体各个部位和组织的过程称为分布。
6. 代谢:药物作为一种异物进入体内后,机体要动员各种机制使药物从体内消除,代谢是药物在体内消除的重要途径。
7. 排泄:药物及其代谢产物主要经尿排泄,其次经粪排泄。
挥发性药物主要经肺随呼出气体排泄。
药物的汗液和乳汁排泄也是药物的排泄途径。
8. 离子障:不论弱酸性或强碱性药物的pKa都是该药在溶液中50%离子化时的PH值,各药有其固定的pKa值。
当与pH的差值以数学值增减时,药物的离子型与非离子型浓度比值以指数值相应变化。
非离子型药物可以自由穿透,而离子型药物就被限制在膜的一侧,这种现象称为离子障。
(书上暂时未找到,来自百度)9. 首关消除:从胃肠道吸收入门静脉系统的药物在到达全身血循环前必先通过肝脏,如果肝脏对其代谢能力很强或由胆汁排泄的量大,则使进入全身血循环内的有效药物量明显减少,这种作用称为首关消除。
(百度)10. 药酶诱导剂:凡能诱导药酶活性增加或加速药酶合成的药物称为药酶诱导剂。
11. 药酶抑制剂:一种能抑制某种酶活性的化学物质。
它通过控制某种酶的活性,来调节或阻抑某些代谢过程,还可抑制耐药性细菌的钝化酶,故可用于某些耐药性细菌感染的治疗。
12. 干肠循环:被分泌到胆汁内的药物及其代谢产物经由胆道及胆总管进入肠腔,然后随粪便排泄出去,经胆汁排入肠腔的药物部分可再经小肠上皮细胞吸收经肝脏进入血液循环,这种肝脏、胆汁、小肠间的循环称干肠循环。
神经递质、受体、激动剂和拮抗剂的类型
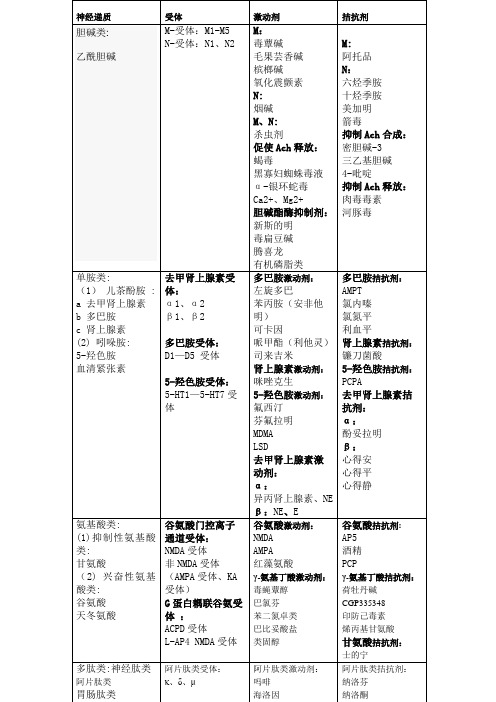
毒蕈碱:在30-60分钟内可出现毒蕈碱样中毒症状。
毒蕈碱样中毒症状是有机磷农药中毒的主要表现.表现为体内多种腺体分泌增加和平滑肌收缩所产生的症状和体征,如多汗,流涎,流泪,鼻溢,和肺部干湿啰音,呼吸困难。
恶心呕吐,腹痛腹泻,肠鸣音亢进,尿频尿急,大小便失禁。
瞳孔缩小,视力模糊,抑制血管平滑肌,血压下降。
毛果芸香碱:对眼和腺体的作用最为明显。
治疗原发性青光眼,包括开角型与闭角型青光眼。
①引起缩瞳,眼压下降,并有调节痉挛等作用。
通过激动瞳孔括约肌的M胆碱受体,使瞳孔括约肌收缩。
缩瞳引起前房角间隙扩大,房水易回流,使眼压下降。
由于睫状肌收缩,悬韧带松弛,使晶状体屈光度增加,故视近物清楚,看远物模糊,称为调节痉挛。
②增加外分泌腺分泌。
对汗腺和唾液腺作用最为明显,尚可增加泪液、胃液、胰液、肠液及呼吸道黏液细胞分泌。
③引起肠道平滑肌兴奋、肌张力增加,支气管平滑肌、尿道、膀胱及胆道肌张力也增加。
槟榔碱在医疗上用于治疗青光眼,能使绦虫瘫痪,所以也用作驱绦虫药,与南瓜仁效果更好。
临床用于治疗产后子宫出血、子宫复旧不良、月经过多等。
烟碱对中枢神经系统尼古丁可与尼古丁乙酰胆碱接受器结合,增加神经传递物的量,脑中的多巴胺增加,产生幸福感和放松感,最后可能会因吸食而有成瘾的现象。
烟草燃烧产生的烟中包含了单胺氧化酶抑制剂(Monoamine oxidase inhibitor),单胺氧化酶会分解单胺类神经传递物、多巴胺、正肾上腺素和血清素。
透过吸烟长期暴露于尼古丁中的人,尼古丁会正向调节小脑和脑干中α4β2尼古丁乙酰胆碱受体。
对周边神经系统尼古丁会刺激交感神经,借由刺激内脏神经影响副肾髓质,释放肾上腺素。
副交感神经节前纤维释放乙酰胆碱,作用在烟碱酸乙酰胆碱接受器上,使之释放肾上腺素和正肾上腺素至血液中。
对副肾髓质尼古丁与肾上腺髓质的烟碱接受器结合后,会增加血液中肾上腺素的含量。
透过与接受器结果,尼古丁使细胞去极化,钙离子由钙离子通道流入,钙离子促使神经细胞以胞泌作用的方式,释出肾上腺素和正肾上腺素至血液中,血液中肾上腺素增加,造成心跳加快,血压升高,呼吸加快,就像高血糖的情形一样。
新型兴奋性氨基酸受体拮抗剂

新型兴奋性氨基酸受体拮抗剂
兴奋性氨基酸(EAA)是哺乳动物脑内的一种内源性神经递质,传递中枢神经系统的大多数兴奋作用。
但EAA及其结构类似物的过量释放对神经元有明显的毒性作用。
过量EAA的持续作用可能导致细胞的急性死亡,而过量EAA的短暂作用可能引起细胞的慢性溃变,细胞的慢性溃变导致各种神经溃变性疾病。
EAA受体拮抗剂能有效地阻断EAA的兴奋毒性作用,因而,从EAA受体拮抗剂中寻找全新的神经系统药物,特别是神经溃变性疾病(如脑缺血性损伤、早老性痴呆、癫痫、巴金森氏症、亨廷顿氏症等)的预防药和治疗药成为近年来神经系统药物研究的主攻方向之一。
根据我们自己的研究成果并参考国外最新进展情况,我们设计了两类新结构类型的化合物。
采用了11条合成路线,共得到40个目的物,其中38个为新化合物。
对所得到的部分目的物,进行了NMDA受体甘氨酸位点的竞争性结合实验,结果显示,所测定的目的物均具有不同程度的NMDA受体甘氨酸位点竞争性拮抗活性,IC50为100~10- ~5μM。
部分目的物还进行了抗脑室注射NMDA引起惊厥的整体动物实验及抗神经细胞缺氧耐受实验,结果也显示所测目的物均具有不同程度的活性。
其余目的物的药理评价正在进行中。
对所有目的物,均在SGI工作站上进行了分子力学计算和分子优势构象的三维图形显示和比较,并进行了初步的构效关系分析。
目前已合成的化合物,不但具有相似于或强于文献报道化合物的拮抗活
性,而且在进一步的结构优化方面有广阔的发展余地,提供了极有意义的先导物。
卫生系统招聘考试《药理学》题库[11-20章]36页
![卫生系统招聘考试《药理学》题库[11-20章]36页](https://img.taocdn.com/s3/m/40ad1d5426284b73f242336c1eb91a37f11132fc.png)
卫生系统招聘考试《药理学》题库[11-20章]36页卫生系统招聘考试《药理学》题库[11-20章]第十一章肾上腺素受体阻断药【内容提示及教材重点】一、α受体阻断药酚妥拉明(phentolamine)作用机制:阻断α1和α2受体药理作用:阻断α受体和直接对血管的作用使血管舒张,用于外周血管痉挛性疾病、嗜铬细胞瘤的诊断和治疗、休克、心肌梗塞和心衰、静滴NA外漏。
二、β受体阻断药普萘洛尔(propranolol)作用机制:阻断β受体。
药理作用:β受体阻断作用,抑制心脏功能;反射性收缩血管;收缩支气管;抑制脂肪分解;抑制肾素释放;内在拟交感活性;膜稳定作用;抑制血小板聚集;减少房水形成。
临床用于心律失常、心绞痛、高血压、甲状腺功能亢进、青光眼。
【作业测试题】一、名词解释1. 肾上腺素升压作用的翻转二、选择题(一) A型题2. 酚妥拉明舒张血管的原理A. 阻断突触后膜α1受体B. 主要奋M受体C. 兴奋突触前膜β受体D. 主要阻断突触前膜α2受体E. 以上均不是3. 普萘洛尔治疗心绞痛的主要药理作用是A. 扩张冠状动脉B. 降低心脏前负荷C. 阻断β受体,减慢心率,抑制心肌收缩力D. 降低左心室壁张力E. 以上都不是4. 下列何药可翻转肾上腺素的升压效应A. 酚妥拉明B. 阿替洛尔C. 利血平D. 阿托品E. 以上都不能5. 治疗外周血管痉挛病可选用A. α受体阻断剂B. α受体激动剂C. β受体阻断剂D. β受体激动剂E. 以上均不行6. 下述哪类药物可诱发或加重支气管哮喘A. α受体激动剂B. α受体阻断剂C. β受体激动剂D. β受体阻断剂E. 以上都不能7. 下面哪种情况,应禁用?受体阻断药A. 心绞痛B. 房室传导阻滞C. 快速型心律失常D. 高血压E. 甲状腺机能亢进8. 普萘洛尔阻断交感神经末梢突触前膜上的β受体,可引起A. 去甲肾上腺素释放减少B. 去甲肾上腺素释放增多C. 心率增加D. 心肌收缩力增强E. 房室传导加快9. 与普萘洛尔有关的禁忌症是A. 青光眼B. 腹胀气C. 尿潴留D. 前列腺肥大E. 支气管哮喘(二) B型题A. 毛果芸香碱B. 利血平C. 敌百虫D. 哌唑嗪E. 阿托品10. 节后拟胆碱药11. M受体阻断药12. 胆碱酯酶抑制剂13. α1受体阻断药14. 影响肾上腺素能神经末梢递质药A. 新斯的明B. 筒箭毒碱C. 去甲肾上腺素D. 异丙肾上腺素E. 酚妥拉明15. 主要兴奋α受体,收缩血管,升高血压16. 能抑制胆碱酯酶,并能直接兴奋N2受体,增强骨骼肌收缩17. 能翻转肾上腺素的升压作用18. 能阻断N2受体,松弛骨骼肌19. 作用于β受体,兴奋心脏,松弛支气管平滑肌(三) X型题20. 能阻断α受体的药物是A. 阿托品B. 氯丙嗪C. 酚妥拉明D. 麦角碱]E. 酚苄明21. β受体阻断药的药理作用是A. 血管收缩冠脉流量减少B. 支气管平滑肌收缩C. 可降眼压D. 心传导减慢E. 心肌抑制,耗氧量降低22. 下列哪些药物对β受体(激动或阻断)有明显的作用A. 普萘洛尔B. 去甲肾上腺素C. 异丙肾上腺素D. 麻黄碱E. 肾上腺素(四) 英文选择题(X型题)23. The following are selective blockers:A. timololB. propanololC. levobunalolD. atenololE. betaxolo24. The following medications can cause conjunctival pigmentation:A. betaxololB. dipivefrineC. azetazolamideD. pilocarpineE. bromonidine25. The following side-effects are seen in b-antagonists:A. reduced tear productionB. decreased corneal sensationC. myopic shiftD. impotenceE. poliosis26. Timolol:A. has a duration of action of between 12 to 24 hoursB. is excreted unchanged by the kidneyC. reduces intraocular pressure because it is b2 selectiveD. can cause dry eyesE. should not be used concurrently as latanoprost27. Propanolol:A. can lower the intraocular pressure if taken orallyB. has a high solubilityC. stabilizes cell membraneD. has partial agonist activityE. can mask hypoglycaemia induced by insulin三. 问答题28. 酚妥拉明的临床用途有哪些?29. 酚苄明与酚妥拉明相比,有哪些特点?30. β受体阻断药的β受体阻断作用包括几方面?31. β受体阻断药物临床禁忌症有哪些?其药理学基础是什么?【参考答案】2. A3. C4. A5. A6. D7. B8. A9. E 10. A 11. E 12. C 13. D 14. B 15. C 16. A 17. E 18. B 19. D 20. BCE 21. BCDE 22. ACDE 23. DE 24. B 25. ABD26. AD 27. ABCE卫生系统招聘考试《药理学》题库[11-20章](2)第十二章局部麻醉药一. 学习要点以适当浓度的药物,局部作用于神经末梢或神经千周围时能暂时、完全和可逆地阻断神经冲动的产生和传导,在意识清醒的条件下,使神经所支配区域的痛觉及运动功能等暂时消失的药物称为局部麻醉药(局麻药)。
蛋白酶激活受体—1阻断剂对兔心肺复苏后脑损伤的保护作用

蛋白酶激活受体—1阻断剂对兔心肺复苏后脑损伤的保护作用心脏骤停后综合征(post-cardiac arrest syndrome,PCAS)是心肺复苏患者自主循环恢复后出现的多器官功能障碍综合征,病死率高、预后差,主要原因是心脏骤停后脑组织持续性损伤[1-2]。
心脏骤停心肺复苏的本质是全身缺血-再灌注损伤,脑组织对缺氧最敏感,因而最易损伤。
心脏骤停后脑损伤虽然由缺血缺氧直接引起,其损伤机制包括兴奋性中毒、钙离子平衡失调、自由基损伤和细胞死亡信号通路活化[3] 。
但是心脏骤停后脑缺血导致的间接损伤未引起足够重视。
近来研究表明,脑缺血或出血时凝血酶可通过受体及非受体介导途径导致脑损伤,其重要途径之一是通过蛋白酶激活受体-1(protease- activated receptor 1,PAR-1)活化加重神经元损伤,其损伤与N-甲基-D-天冬氨酸受体(NMDA受体)有关。
PAR-1激活可上调NMDA受体介导的兴奋性氨基酸毒性,从而导致细胞凋亡[4]。
此外,凝血酶活化PAR-1,引起脑组织炎症反应,亦是脑损伤的重要原因。
因此,阻断凝血酶激活PAR-1应该是防治脑损伤的有效途径。
本研究应用兔心脏骤停后综合征模型,探索应用蛋白酶激活受体-1的特异性阻断剂对心肺复苏后脑损伤的影响。
1 材料与方法1.1 实验动物与模型建立本动物实验得到湖北医药学院动物实验伦理委员会的批准。
日本大耳白兔30只(湖北省实验动物研究中心提供)6月龄,雄性,体质量(2.43±0.23)kg,随机(随机数字法)分为假手术组、心脏骤停后综合征(PCAS)组、PAR-1阻断剂组,每组10只。
兔心脏骤停复苏后综合征(PCAS)制备方法依据文献[5]方法进行,简述如下。
兔速眠新0.24 mL/kg麻醉后固定,用24号留置针穿刺股动脉,连接ZOLL全功能除颤监护起搏仪,进行心电监护。
逆行气管插管,固定气管导管。
心率、呼吸、血压稳定5 min后(以此作为实验计时点,即0时),采集动物血液,-80 ℃储藏供检测用。
2021执业药师继续教育阿尔茨海默病及治疗答案

2021执业药师继续教育阿尔茨海默病及治疗答案1 . (单选题)老年痴呆是一种中枢神经系统变性病,主要表现不包括()A .渐进性记忆障碍B .认知功能障碍C .人格改变D .出现帕金森2 . (单选题)健忘和老年痴呆有本质的区别,健忘表现不包括()A .脑发生病理性变化,器质性的智能衰退B .虽会记错日期,有时讲前忘后,但他们仍能料理自己的生活,甚至能照顾家人C .有七情六欲D .对做过事情的遗忘总是部分性的3 . (单选题)应该进行老年痴呆检查的人群不包括()A .65岁以上的老年人B .自觉或者家属发现有记忆力下降者C .肥胖D .家族中有老年痴呆的患者4 . (单选题)简易精神状态检查(MMSE)量表评分标准为正常的是()A .受教育年限大于等于7年者,>24分为正常B .受教育年限大于等于7年者,>26分为正常C .受教育年限大于等于7年者,>27分为正常D .受教育年限大于等于7年者,>28分为正常5 . (单选题)治疗老年痴呆的药物中,属于兴奋性氨基酸受体拮抗剂的是()A .卡巴拉汀B .多奈哌齐C .美金刚D .甘露特钠6 . (单选题)多奈哌齐每日最大剂量不得超过()A .5mgB .10mgC .15mgD .20mg7 . (单选题)卡巴拉汀每日最大剂量不得超过()A .3mgB .6mgC .9mD .12mg8 . (单选题)美金刚每日最大剂量不得超过()A .5mgB .10mgC .15mgD .20mg9 . (单选题)甘露特钠治疗老年痴呆的作用机理不包括()A .抑制Aβ斑块形成B .调节脂质代谢C .平衡肠道菌群D .减轻神经炎症10 . (单选题)预防老年痴呆,需要在身体上注意的方面不包括()A .防治心脏病B .控制血压和血糖C .保护大脑D .口腔检查。
营养神经药物

GABA受体激动剂GABA是脑内主要的抑制性神经递质,它的作用在于兴奋性氨基酸递质谷氨酸起平衡作用。
GABA受体激活后能抑制兴奋性神经毒作用,如muscimol、MK-801、clomethiaiole等。
伽马氨基丁酸是中枢神经系统中很重要的抑制性神经递质,它是一种天然存在的非蛋白组成氨基酸,具有极其重要的生理功能,它能促进脑的活化性,健脑益智,抗癫痫,促进睡眠,美容润肤,延缓脑衰老机能,能补充人体抑制性神经递质,具有良好的降血压功效。
促进肾机能改善和保护作用。
抑制脂肪肝及肥胖症,活化肝功能。
每日补充微量的伽玛氨基丁酸有利于心脑血压的缓解,又能促进人体内氨基酸代谢的平衡,调节免疫功能。
抗氧化剂、自由基清除和其它:维生素E、维生素C和甘露醇也具有抗氧化和自由基清除的作用。
缺血缺氧导致脑组织发生一系列还原反应其中脂质产生的氧自由基是在灌注损害的重要原因,动物实验中抗氧化剂tirilazad、ebselen 能减少严重梗死动物模型的梗死容积,其他具有潜在作用是药物如超氧化物歧化酶(SOD)、谷胱甘肽、甘露醇等都有抗自由基作用。
白细胞黏附抑制剂脑缺血后,在受损部位的白细胞浸润和炎性细胞因子参与了缺血诱发的组织损伤过程,其中白细胞介素-1β和肿瘤坏死因子-а的过量产生加重脑缺血损害,而转化生长因子-β1的产生对脑缺血具有保护作用。
动物实验表明,多种抗白细胞黏附的抗体可减轻脑缺血时的神经损伤。
营养神经药物有纳络酮针、奥拉西坦针、鼠神经生长因子针等,另外还有神经保护剂如依达拉奉针等。
艾地苯醌是改善脑代谢的,也可以说是营养脑细胞的。
脑缺血后可诱导许多神经保护因子的表达,产生的神经营养因子(NTF)、神经生长因子(NGF)、脑源性神经营养因子(BDNF)、硷性成纤维细胞生长因子(bFGF)、胰岛素样生长因子(IGF)、转化生长因子(TGF)等都对神经元起保护和营养作用。
外源性神经营养因子类药物在细胞培养中效果明显,在动物模型和临床上效果不佳,可能与局部药物浓度及其他因素有关。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
兴奋性氨基酸(Excitatory amino acid,EAA)是广泛存在于哺乳类动物中枢神经系统的正常兴奋性神经递质,参与突触兴奋传递,学习记忆形成以及与多种神经变性疾病有关。
缺血、缺氧、创伤、中毒等因素能触发中枢神经系统的EAA过度兴奋,在能量代谢失衡的基础上,异常堆积,产生神经毒性作用。
急性颅脑损伤后脑内EAA的浓度变化与脑损伤的程度有关。
本文综述了EAA的生物学基础、毒理作用机制、影响因素,及对神经系统的损伤机制和近年来国内外的研究现状。
1 兴奋性氨基酸及其受体
兴奋性氨基酸是广泛存在于哺乳类动物中枢神经系统的正常兴奋性神经递质,参与突触兴奋传递,学习记忆形成以及与多种神经变性疾病有关。
在胚胎时期海马神经上皮中即存在向海马结构中分化的兴奋性氨基酸前体细胞,随着其层次结构出现,这些前体细胞不断分化、迁移并定居于各层之间,尤以锥体层和颗粒层最多。
EAA包括谷氨酸(Glu)、天冬氨酸(Asp)、N-甲基-D-天冬氨酸(NMDA)、亮氨酸等。
其中谷氨酸(Glu)是中枢神经系统内含量最高的一种氨基酸,Glu在中枢神经系统的分布不均,以大脑皮层、小脑、纹状体的含量最高,脑干、下丘脑次之。
脊髓中含量明显低于脑干,但有其特异分布,背根含量高于腹根,背侧灰质含量高于腹侧灰质。
因此,有人推测Glu为初级感觉传入纤维的兴奋性递质。
NMDA是通过改变Asp
的结构而合成的EAA,兴奋作用可达Glu的100倍。
目前已知的EAA受体可分为两大类:NMDA受体和非NMDA受体。
非NMDA受体包括AMPA(α-氨基-3-羟基-5-甲基-4-异唑丙酸)受体、KA(海人藻酸)受体、QA(使君子酸)受体、代谢型受体和L-AP4受体等。
NMDA和AMPA为离子型受体,主要作用于Ca 2+ 、Na + 通道。
NMDA受体对Ca 2+ 通道的开放受甘氨酸的抑制性调节,并可被Mg 2+ 以电压依赖方式阻断。
KA、QA受体开放单价离子通道,允许Na + 、Cl - 、H 2 O进入细胞内,促使神经元肿胀坏死。
代谢型受体通过G - 蛋白偶联,调节细胞膜离子通道和酶活性,与神经元的异常放电有关。
有研究发现[9],NMDA受体主要介导皮肤的伤害性传入,非NMDA受体主要介导肌肉和内脏的伤害性传入。
皮肤和肌肉的伤害性传入分别诱发释放更多的门冬氨酸和谷氨酸可能是这种差别的主要原因之一。
NMDA受体的不同调节位点在伤害性信息传递中有密切的协同作用。
2 兴奋性氨基酸的毒理作用机制
正常情况下,EAA主要存在于神经末梢的突触囊泡内,末梢去极化时释放到突触间隙,作用于突触后膜的特异受体,完成兴奋性突触传递及其它生理作用。
释放入突触间隙的EAA除刺激突触后膜的EA A受体发挥生理效应外,绝大部分可经EAA能神经末梢重摄取而回收,少部分可被胶质细胞所摄取。
EAA的重摄取主要依靠突触前膜上的EAA转运体来完成,此转运体是Na + 依赖性的,即转运过程必须依赖细胞外的Na + 。
缺血、缺氧、创伤、中毒等因素触发中枢神经系统的EAA过度兴奋,在能量代谢失衡的基础上,异常堆积,促
进具有细胞毒性的反应性氮中间产物(RNI)、反应性氧中间产物(R OI)形成,促使细胞外的Ca 2+ 大量内流,激活磷酸酯酶、蛋白酶、一氧化氮合酶等,引起神经细胞的坏死或凋亡。
EAAs造成神经毒性作用主要有两个过程,一是作用于非NMDA受体,引起Na + 、Cl - 、H 2 O的内流,造成神经元急性水肿为特征的急性损伤过程,此过程在刺激因素去除后可以恢复。
二是通过激兴奋NMDA受体,直接或间接启动电压依赖性通道,Ca 2+ 大量内流,造成迟发性的神经元变性坏死的迟发过程,该过程为不可逆。
3 兴奋性氨基酸导致的神经系统的损伤
Castillo等在128例急性缺血性卒中患者的EAA检测中,发现血浆Glu、Asp含量在急性期明显升高,并与脑梗死体积和病情轻重正相关。
缺血性脑卒中的发病过程中,在能量耗竭的缺血中心,EAA的毒性引起神经元坏死;在能量相对缺乏的半暗带区,则导致神经细胞凋亡。
线粒体功能的损伤程度决定了神经元的死亡方式。
李燕珍等用高效液相色谱法测定42例出血性脑血管病患者血清证实Glu、Asp含量在急性期(1~5天)显著高于对照组,且与出血量呈正比。
用流体冲击装置制作中度颅脑损伤模型的大鼠,发现脑损伤后脑血流明显下降,脑组织内Glu、Asp明显升高。
原因可能是局部缺血,能量耗竭,膜通透性改变导致K + 外流,引起神经去极化和神经兴奋冲动发放,促使Glu和Asp释放[11]。
脑缺血时,胞外Glu浓度变化强烈地影响着缺血神经细胞损伤,脑缺血引起神经细胞损伤易发部位与EAA 在脑内的分布一致,给予NMDA受体拮抗剂可减少神经细胞损伤。
急
性颅脑损伤后脑内兴奋性氨基酸的浓度变化与脑损伤的程度有关,损伤(根据格拉斯哥昏迷分级计分)越重,升高越明显。
赵宝东等通过实验证实给离体培养的大鼠小脑神经元培养液中添加EAA后24h,可以引起细胞的急性损伤作用,导致细胞的贴壁能力下降,突起减少;
台盼兰染色见死亡细胞明显增多,与对照组存在显著差异。
黄文等向生长良好的体外培养神经元添加Glu后,发现神经元逐渐退化,胞体肿胀,突起断裂,神经元崩解。
4 影响兴奋性氨基酸作用的因素
EAA对神经细胞的毒性作用受多种因素的影响。
赵宝东等证实N GF能降低急性全脑缺血再灌注期间兴奋性氨基酸的释放。
邱小鹰等发现脑缺血时,轻度高温(38.8~39.4℃)组Glu、Asp含量明显高于常温(36.5~37℃)组或假手术组(P<0.01);亚低温(32~32.5℃)组明显低于常温组或轻度高温组(P<0.01);再灌注后3h内,各脑温组Glu、Asp均有不同程度下降,说明脑温对EAA的释放具有一定的调控作用。
脑温变化也可改变缺血神经细胞的损伤结局,高温可加重缺血神经细胞损伤、加快缺血“半暗带”区向梗死区演化、增加脑梗死体积;亚低温可减轻缺血神经细胞损伤、抑制缺血“半暗带”区向梗死区演化、减少脑梗死体积,且可能与脑温对脑缺血后胞外Glu浓度的影响有关。
腹侧压迫损伤法造成大鼠脊髓中度损伤,伤后立即灌注非特异性腺苷受体激动剂2-氯腺苷(2-CADO,0.1mmol/L)。
发现2-CA DO可显著抑制Glu,Asp的升高(P<0.01)。
应用硫酸镁治疗脑梗塞后EAA含量明显降低,表明硫酸镁可作为EAA的阻断剂,能降低
EAA的神经毒性反应。
其作用机制可能为:(1)阻断电压依赖性NM DA离子通道;(2)阻断钙离子内流,防止细胞内钙超载;(3)硫酸镁有保护细胞膜的功能。
实验证实:外源性Glu迅速激活了脊髓组织一氧化氮合酶(NOS)的活性,而抑制NOS活性则明显降低了伤段脊髓组织Glu的浓度。
由此可见脊髓伤后脊髓组织中NOS与EAA的释放相互促进,因而在继发性脊髓损伤中形成级联放大的神经毒性因子释放。
研究发现654-2可明显减轻缺血缺氧新生鼠脑组织Glu的过度产生,表明654-2对缺血缺氧新生大鼠脑损伤有一定的保护作用。
电刺激丘脑底核后,改善了帕金森病大鼠的旋转行为,纹状体中Glu含量明显减少(P<0.01),Asp则无明显改变(P>0.05),可见高频电刺激丘脑底核抑制了同侧谷氨酸的释放。
用放射免疫法和高效液相色谱电化学检测器分别测定了SAMP8小鼠血浆、大脑皮层、海马、下丘脑皮质酮(corticosterone,CORT)水平和大脑皮层、海马EAA含量。
结果显示,SAMP8小鼠血浆CORT水平均显著高于其同龄正常对照小鼠SAMR1(P<0.05,或P<0.01)。
SAMP8小鼠8月和12月龄大脑皮层和海马CORT水平也明显高于同龄正常对照SAMR1小鼠(P<0.05,或P<0.01)。
同时,SAMP8小鼠大脑皮层和海马Glu 含量有上升趋势,谷氨酰胺(Gln)含量则明显高于SAMR1小鼠。
S AMP8小鼠切除肾上腺后,大脑皮层和海马Glu含量呈下降趋势,而Gln含量则显著降低。
给切除肾上腺的SAMP8小鼠补充CORT(ip,10mg/kg),大脑皮层和海马Glu和Gln水平均明显升高(P<0.05,或P<0.01),而Asp含量始终没有变化。
结果提示,SAMP8小鼠大
脑皮层和海马Glu和Gln水平升高是肾上腺依赖的。
SAMP8小鼠大脑皮层和海马Glu和Gln含量升高可能与其大脑皮层和海马CORT 水平升高有关。
5 存在问题及展望
(1)目前对EAA的研究对象局限于脑出血、脑缺血以及癫痫的机体,除裴发光等对儿童氟乙酰胺中毒所致的EAA变化外,对其他中毒条件下造成的神经损伤研究甚少。
故对中毒机体的EAAS变化的研究成为毒理工作者的当务之急。
(2)目前对EAA的研究基本局限于在体动物研究,由于受血流状态影响以及神经体液的调节作用,其结果尚不能说明特定神经元的特征性改变。
故应通过体外细胞(组织)培养的方法,研究EAA对特定细胞(核团)的作用机制。