DNA的重组与转座
分子生物学:重组和转座

转座子的特点
转座子是不必借助同源序列就可以移动的片断,即转座作用与供体 和受体的序列无关;
原核生物和真核生物都有转座子; 转座序列可沿染色体移动,甚至在不同染色体间跳跃(跳跃基因) 转座子对基因组而言是一个不稳定因素,它可导致宿主序列删除、
倒位或易位,并且其在基因组中成为“可移动的同源区”。位于 不同位点的两个拷贝转座子之间可以发生交互重组,从而造成基 因组不同形式的重排。 有些转座子与基因组的关系犹如寄生,它们的功能只是为了自身 的扩增与繁衍,因此被称为是自私的DNA。
转座中涉及的机制依赖于DNA 链的切割和重接,因此 (branch migration)
Cro蛋白抑制C I基因的转录,它占优势噬菌体即进入繁殖周期,并导致宿主细胞裂解。
与重组过程联系起来。 二者含有共同的核心序列15bp(O区)。
DNA转座子上携带作为重组位点的DNA序列,以及参与重组的蛋白质的基因(转座酶基因)。 同源重组 (homologous recombination )或普遍性重组(generalized recombination )
复制式转座 如 Tn3
非复制式转座 如 Tn10
类病毒反转录转座子/反转录病毒
带有反向终端重复序列.反向终端 重复序列嵌入在较长的正向排列的 重复序列(长末端重复序列, long terminal repeats LTR)中;
带有靶位点重复序列; 类病毒反转录转座子编码两种移位所需的蛋白:转座酶和反转录酶。 类病毒反转录转座子和反转录病毒的区别在于:反转录病毒的基因
径的歧化做好准备。
位点特异性重组与同源重组的区别
带有2个基因ORF1和ORF2;
同源重组 (homologous recombination )或普遍性重组(generalized recombination )
第二十三章重组和转座
第七节 真核生物的转座因子
一、玉米中的控制因子
1938年Marcus Rhoades首次发现不稳定突变等
位基因(unstable mutant allele),即一种回 复突变率很高的等位基因。不稳定是取 决于不连锁的Dt基因的存在。
McClintock。1940~1950描述了大量的控制因子
A1 A1 dt dt
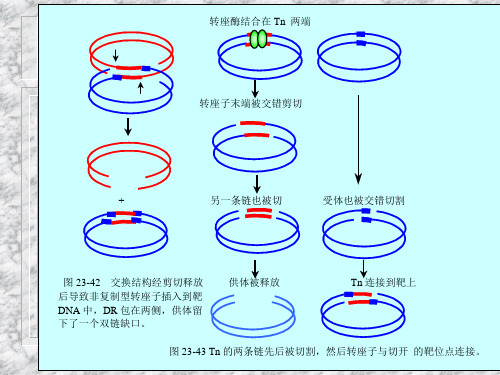
转座酶结合在 Tn 两端 转座子末端被交错剪切
+
另一条链也被切
受体也被交错切割
图 23-42 交换结构经剪切释放 后导致非复制型转座子插入到靶 DNA 中,DR 包在两侧,供体留 下了一个双链缺口。
供体被释放
Tn 连接到靶上
图 23-43 Tn 的两条链先后被切割,然后转座子与切开 的靶位点连接。
复制型转座模型解释了 (1) 复制性转座在转座后原来的位置上保留原
LTR 反转录病毒 gag
LTR pol int env
δ gag int
pol
δ
Ty
TyA
TyB
LTR gag int
pol LTR
Copia
ORF
pol
L1
ORF1
ORF2
图 23-72 病毒家族的反转录子末端有重复顺序内部有 开放读框
无LTR的反转录转 座子通过切开靶位 点双链,提供了引 物末端。反转录转 座子作为模板合成
Copia(果蝇)
Pol Ⅲ 转录 的加 工假 基
因
LINES L1(哺乳动物)
末端
长末端重复序列
无末端重复序列
靶重复序列
4~6 bp
7~12bp
阅读框
反转录酶和/或整合酶 无(不编码和转座有关
分子生物学第7章 DNA的重组与转座

7.2.1 λ phage DNA的整合与切除 1、实现机制:
均是通过---细菌DNA和λDNA上特定位点之间的重组
2、特定位点-----附着位点(attachment site att)
◘ 整合过程需要λ整合酶 (integrase Int)(λ编码)
和寄主的整合宿主因子IHF (integration host factor) 共同作用
◘ 整合后的附着位点为 attL(BOP’) attR(POB’)
溶菌周期 (lysis)
溶源性细菌 原噬菌体 (lysogen)
4、整合分子机制
5’
5’
b
3’
酶切
(8)
A
B
两臂旋转
b a
(9)
A
B
(3) 5’ A 3’ 3’ 5’ a
(4) 5’ A 3’ 3’ 5’ a
(5) 5’ A 3’ 3’ 5’ a
(6) 5’ A 3’ 3’ 5’ a
(7)
A
B 3’
5’
b
5’
b
3’
游离端移动联会
B 3’
(10)
A
5’
B
5’
b 3’ 游离端交叉连接
一、位点特异性重组( site-specific recombination) 1、概念:发生在专一序列而顺序极少相同的DNA分子间 的重组
噬菌体基因组整合到细菌染色体基因组中属此种重组
2、特征:
在特定的结合序列部位,有专一的酶催化断裂重接---产生精确的DNA 重排 都具有整合作用的两个基本特征
第二十三章 DNA重组和重组DNA技术【生物化学与分子生物学 9版原版】

第二节
重组DNA技术
序言:
重组DNA技术
又称: 分子克隆(molecular cloning) DNA克隆(DNA cloning) 基因工程(genetic engineering)
主要过程:
——在体外将目的DNA与能自主复制的遗传元件(载体)连接, 形成重组DNA分子 ——重组DNA分子在受体细胞中复制、扩增及克隆化,从而获得 单一DNA分子的大量拷贝
• 大多数RE的识别序列为回文结构(palindrome) • 有些RE所识别的序列虽然不完全相同,但切割DNA双链后可产
生相同的黏端,这样的酶彼此互称同尾酶(isocaudamer) • 有些RE虽然来源不同,但能识别同一序列(切割位点可相同或
不同),这样的两种酶成同裂酶(isoschizomer)
克隆载体
——是指用于外源DNA片段的克隆和在受体细胞中扩增的DNA分子。
克隆载体的基本特点:
至少有一个复制起点使载体能在宿主细胞中自主复制,并能使克隆的外源DNA片段得到 同步扩增; 至少有一个选择标志(selection marker),从而区分含有载体和不含有载体的细胞,如 抗生素抗性基因、-半乳糖苷酶基因(lacZ)、营养缺陷耐受基因等; 有适宜的RE单一切点,可供外源基因插入载体。
第一节小结
小结:
自然界DNA重组方式 主要包括: 同源重组,Holliday模式是最经典的同源重组模式 位点特异性重组 转座重组或转座,包括插入序列和转座子的重组 接合,通过细胞接触所发生的基因转移 转化,通过细胞自主摄取发生的DNA整合 转导,病毒感染介导的DNA整合 CRISPR/Cas9系统,细菌获得病毒DNA用于攻击病毒
一、同源重组
同源重组的Holliday模型
分子生学技术-第七章 DNA的重组与转座

2、复合式转座子
它是一类较复杂的转座子,由两个重复序列 夹着一个或多个结构基因组成。除了含有转位相 关的功能外,还带有一些其他基因,如抗药性基 因、抗重金属离子基因等。 它们都具有末端反向重复序列、可编码基因 和插入位点的正向重复序列
TnA转座子:
结构较复杂,长度约5000bp。两端没 有IS结构,仅含有两个末端反向重复序列, 一般约38bp。内部通常含有转位相关的基 因和抗药性基因等,插入位点约5bp。
②果蝇中的转座子—P转座子 两端有31bp的反向重复序列,靶位点产生8bp 的正向重复序列,有四个外显子和三个内含子, 在体细胞中,只有前两个内含子被切除,产生一个 转座阻遏蛋白;在卵细胞中,三个内含子全部被切 除,可产生转座酶,导致P转座子转座和后代不育。
四、转座的遗传学效应
a、转座引起插入突变 各种IS、Tn转座子都可引起突变 b、转座产生新的基因 转座上存在一些抗药性基因或抗重金属基因 等
一、霍利迪结构(Holliday Structure)
霍利迪结构:它是DNA重组过程中的一个中间体, 重组时两个DNA双链体以四股DNA在连结点形成的十字 形DNA分子构象。
异源双链 (Hetero duplex):指在重组部位,每条双 链中均有一段DNA链来自另一个双链中的相对链。
分支迁移(Branch migration):一旦两个重组的 DNA分子形成霍利迪结构,交叉连接点很容易像拉链一 样移动,从而扩大异源双链体区域,这一过程叫分支迁 移。
2、Rec BCD核酸酶 Rec BCD蛋白具有DNA解螺旋酶、核 酸内切酶和外切酶的活性。它可以帮助有游 离末端的DNA形成单链DNA片段,从而有 利于RecA 蛋白作用。 它识别并切割Chi序列:5'GCTGGTGG-3'
3 遗传重组与转座(第3节至第4节)

双转座子插入所引起的外显子改组示意图
8、真核生物的转座成分
根据转座机制目前分为两类: a) 转座机制与细菌的转座子类似 遗传信息: DNA→DNA
♥ 玉米的Ac-Ds元件、果蝇的P元件和FB元件等
b) 转作机制类似逆转录病毒 遗传信息: RNA→DNA→RNA
♥
如:逆转录病毒、果蝇的Copia元件、酵母的Ty元件
不准确切除:留下转座子残迹,产生插入突变,但 转座子标志消失。
转座子切离所造成的序列变异
⑥外显子改组
当二个转座子被同一转座酶识别而整合到染色体的邻
近位置时,则位于它们之间的序列有可能被转座酶作用而 转座,如果这DNA序列中含有外显子,则被切离并可能 插入另一基因中,这种效应称为外显子改组(exon shuffling)( 图)。 外显子改组将导致基因组中新基因的产生。
得1983年的诺贝尔奖。
玉米地中的先知 Barbara McClintock (芭芭拉· 麦克林托克):1902-1992
2
转座子的定义
1)转座子(元)或转座元件 (transposon or transposable element): 即能够反复插入到基因中许多位点的特殊DNA片段, 它们可从一个位点转移到另一个位点,从一个复制子到另 一个复制子。
M型(母本贡献的,maternal contributing)
M(♂)×P(♀) P(♂)×M(♀) 后代不育 后代可育。
阻遏P因子的转座
转座酶
雄性染色体 P 因子
ORF0 ORF1 ORF2 ORF3
P 品系 (P♂×P♀) 雌性染色体 P 因子
ORF0 ORF1 ORF2 ORF3
P 细胞型 66KD 阻遏物
分子生物学—同源重组、位点专一性重组、转座

分⼦⽣物学—同源重组、位点专⼀性重组、转座同源重组遗传交换、染⾊体上基因重排、断裂DNA的修复(1)同源重组的形式常发⽣在同源染⾊体之间(分⼦间重组)也可发⽣在同⼀DNA分⼦内(分⼦内重组)(2)Holliday 模型解释同源重组的⼀个经典模型基于重组过程中有⼗字形的中间产物(3)RecBCD重组途径存于E. coli中,需要RecBCD蛋⽩(RecBCD protein,recB、recC、recD基因的产物)参与DNA损伤造成的双链断裂以及外源DNA线性分⼦的DNA断端。
RecBCD是⼀种具多功能的酶,依赖于DNA的ATPase(⽔解ATP, 为DNA的解螺旋提供能量);DNA helicase; DNA nuclease,可作⽤于单链或双链DNA, 切割χ位点(GCTGGTGG)χ(chi): crossover hotspot instigator RecBCD的切点与底物的序列有关,可在χ位点3’端4 ~ 6 ntRecA :38 kD 的蛋⽩质,具多种功能,在重组中促进同源DNA单链的交换(主要是⼀单链与⼀双链中的同源单链交换);交换过程需ATP。
RuvA和RuvB :RuvA和RuvB均具DNA helicase活性,RuvB还有ATPase活性;RuvA特异性识别并结合到Holliday junction,并促使RuvB蛋⽩结合到这⼀位点;RuvB⽔解ATP,提供能量,促使分⽀迁移RuvC :解开Holliday junction的核酸内切酶,特异地与Holliday junction结合,在特异位点切开Holliday junction。
RuvC是⼆聚体,有2个活性位点,能切开Holliday junction的两条链,切割位点有特异的序列,必须等branch migration 到达特异序列后,RuvC才能起作⽤,以何种⽅式解开Holliday junction取决于RuvC切割位点在两对同源单链上的频率。
DNA重组的机理---精华版

异常分离与基因转变
在一个杂合体中,如果一染色体把基因A交给它的 同源染色体,则它的同源染色体必定把基因a交回给 它,所以在真菌中,一个座位上的两等位基因分离时 ,应该呈现2:2或1:1:1:1或1:2:1的分离(表8-1) ,这就说明重组通常总是交互的。可是Lindegren在面 包酵母(Saccharomyces cerevisiae)中发现,有的子囊 含有(3A+1a)或(1A+3a)的子囊孢子。以后在脉 孢霉、酿酒酵母、子囊菌Ascobolus immersus及果蝇中 也发现这种现象。
•片段重组体:切开的链与原来断裂 的是同一条链,重组体含有一段异源 双链区,其两侧来自同一亲本DNA。 •拼接重组体:切开的链并非原来断 裂的链,重组体异源双链区的两侧来 自不同亲本DNA。
电镜下的Holliday结构
Meselson-Radding单链侵入模型
Holliday模型中为对称的杂合双链, 而实际情况有不均等分离现象,1975年 Meselson-Radding 提出模型解释这种不 对称重组现象。
基因重组的相关概念 同源重组 转座重组 位点特异性重组 DNA重组技术简介
一、基因重组
• • 英文名称:gene recombination 定义:是指由于不同DNA链的断裂和连 接而产生的DNA片段的交换、插入等的 重新组合,形成新的DNA分子的过程。 包括发生在生物体内(如减数分裂中异源 双链的核酸交换)和在体外环境中用人工 手段使不同来源DNA重新组合的过程。
真核生物的转座子
Barbara McClintock
Nobel Prize for Physiology Medicine 1983
玉米的Ac-Ds系统
真核生物的转座子根据其DNA结构和转座 机理的不同,分为两类,第一类是反转座子,其 转座过程是DNA→RNA→DNA,需要RNA中 间体;第二类是DNA转座子,其转座过程是 DNA→DNA,无RNA中间体。
DNA的重组(考研分子)知识点梳理

DNA的重组(考研分子)知识点梳理1.D NA分子内或分子间发生遗传信息的重新组合称为遗传重组(genetic recombination)或基因重排(genetic rearrangement),重组产物为重组体DNA,DNA的重组广泛存在于各位生物中,真核生物多发生在减数分裂同源染色体之间的交换2.D NA重组能够迅速增加群体的遗传多样性,使有利的变异与有害的变异分开,通过优化组合积累有意义的遗传信息;为DNA的损伤或复制障碍提供修复机制;调控某些基因的表达或生物的发育过程3.D NA重组(recombination)●概念:指发生在DNA分子内或DNA分子之间的核苷酸序列的交换、重排和转移的现象,是已有遗传物质的重新组合过程,包括同源重组、位点特异性重组和转座重组,是基因变异和物种进化的遗传基础●作用:生物体利用重组产生新的基因或等位基因组合、病毒利用重组将自身DNA整合到宿主细胞DNA中;基因工程技术利用重组进行基因敲除和遗传作图、生物体利用重组进行重组修复●分类●同源重组(homologous recombination)●概念:又称为一般性重组(general recombination),它是由两条具有同源区的DNA分子,通过配对、链的断裂和再连接而产生片段交换的过程,不依赖序列的特异性,只依赖序列的同源性,包括细菌的接合、转化和转导真核生物同源染色体之间的交换等●同源重组的类型●解释同源重组机制的模型●Holliday 模型●内容1)两个同源染色体DNA相互靠近,排列整齐2)两条链在对应的位置上各产生一个单链的断裂3)被切开的链相互入侵、交换、连接形成Holliday中间体4)通过分支迁移产生异源双链DNA5)Holliday中间体切开并修复,形成两个双链重组DNA,根据切开方向分为片段重组体(切开的链与原断裂的链为同一条,产生的重组体有一段异源双链区,异源双链区两侧来自同一亲本DNA链,左)和剪接重组体(切开的链不是原来断裂的链,重组体异源双链区两侧不是来自同一亲本DNA,右)●不足:Holliday模型能够较好地解释同源重组现象,但是两个DNA 分子对应链的相同位置发生断裂的可能性很小●单链断裂模型1)两个同源染色体中某一个DNA分子的一条链发生断裂产生3'末端,入侵另一个DNA分子的同源区的互补链并与之配对,●双链断裂模型1)在一个DNA分子中产生一个双链断裂,DNA外切酶在断裂处进行修剪,产生3'末端,3'末端入侵另一条DNA分子并与之配对,对应的链则被置换出来以原来断裂的链配对,经过修复合成和链的再连接形成两个交叉●细菌的基因转移与重组的机制●接合(conjugation)1)细菌的细胞相互接触时遗传信息从一个细胞转移到另一个细胞的过程2)供体细胞被定义为雄性,受体细胞被定义为雌性,转移DNA的过程由接合质粒完成,能促使染色体DNA基因转移的接合质粒称为F因子,F因子中与基因转移有关的转移区约占整个染色体的三分之一,其中的tra基因编码的蛋白质是构成F性菌毛的亚基,F性菌毛接触受体细胞表面后接合过程被活化,tra S和traT基因编码表面排斥蛋白(防止与F⁺细菌接合),F⁺细菌与受体细胞靠近后,TraD蛋白(一种内膜蛋白)作为DNA的转移通道,Tra I在Tra Y的协助下结合到接合质粒的转移起点切开一条链并与5'端共价结合,Tra I也有解旋酶活性,5'端进入受体细胞内便开始合成互补链,因此受体细胞变为F⁺细胞,而供体中的单链也合成互补链3)F因子也能整合在大肠杆菌染色体DNA上,当F因子启动接合时,质粒基因中转移起点被切开,其前导链引导染色体DNA单链转移,至于转移DNA链长短取决于转移过程的时间,转移到受体中的DNA单链先互补配对成双链,然后与受体DNA发生重组,外源基因的插入需要在两端分别形成交叉连接,即发生两个位点的重组4)整合在染色体DNA中的F因子也可能被切下来,不精确切割使切下来的F因子常带有宿主的染色体基因,称为F'因子,F'细胞与F⁻细胞杂交,供体部分的染色体基因随F'因子进入受体细胞,无需整合就能表达(因为接合质粒上有转录起始点),实际上形成部分二聚体,此时受体细胞变为F',该过程叫性导(sexduction)●遗传转化(genetic transformation)1)是指细菌品系由于吸收了外源DNA(转化因子)而发生遗传性状改变的现象,具有摄取周围环境中游离DNA分子能力的细菌细胞称为感受态细胞(competence cell),感受态是暂时的,与特定的生理状态有关2)感受态因子可以诱导与感受态有关的蛋白的表达,如自溶素能使细胞表面的DNA结合蛋白和核酸酶暴露,当游离的DNA与DNA结合蛋白结合后,核酸酶使其一条链降解,另一条链被吸收进入细胞,并与感受态特异蛋白结合整合到染色体DNA中发生重组3)也有少数细菌可以吸收双链DNA4)用高浓度Ca²⁺处理大肠杆菌可以使其转化为感受态●转导(transduction)1)是通过噬菌体将细菌基因从供体转移到受体细胞的过程,包括普遍性转导和局限性转导2)普遍性转导:宿主细胞基因组任意一段DNA组装到成熟噬菌体颗粒内而被代入受体菌3)局限性转导:某些温和的噬菌体在装配病毒颗粒时将宿主的染色体整合部分的DNA切割下来取代病毒DNA●细胞融合(cell fusion)1)由于细菌细胞质膜融合导致基因转移和重组,实验室内可以通过使用溶菌酶将去除细菌细胞壁的肽聚糖使之成为原生质体进而完成融合●与重组有关的酶1)RecA蛋白●此蛋白可以诱发SOS反应和促进DNA单链的同化(单链与同源DNA分子双链发生链的交换)从而使重组过程中DNA配对、形成Holliday中间体和分支移动等过程能够实现●机理:数千个RecA蛋白单体与DNA单链结合形成螺旋状纤丝,此复合物与双链DNA作用使其部分解旋,然后迅速扫描单链DNA的互补序列,一旦找到,互补区双链进一步解旋与单链DNA沿5'——→3'方向互补配对,直到交换终止(ATP提供能量)●该基因突变会导致双链断裂积累,无法形成正常的联会复合体●真核生物中RecA的类似物:酵母中的Rad51和Dmc1蛋白;人体中的BCRA1和BCRA2蛋白,若编码这俩蛋白的基因突变会导致乳腺癌2)RecBCD蛋白●多功能酶:依赖于ATP的核酸外切酶活性、可被ATP增强的核酸内切酶活性(内外切都有)、ATP依赖的解旋酶活性●DNA分子发生双链断裂后,RecBCD结合在游离端,使DNA双链解旋并降解,当其移动到χ 位点(5'-GCTGGTGG-3'),在其3'端4—6个核苷酸处切开,产生有3'端的DNA单链,之后结合有RecA和SSB的DNA单链入侵双链DNA形成D环结构,RecA协助寻找同源区,互补配对后RecA和SSB被释放3)Ruv A、Ruv B和Ruv C蛋白●Ruv A蛋白能够识别Holliday 连结体的交叉点并结合,使其变成四方平面构象,并帮助Ruv B蛋白六聚体在交叉点上游结合在DNA双链上,通过水解ATP使双链解旋,并使异源链螺旋化,最后Ruv C蛋白切开联结体(分支迁移就是为了找到ATTG序列,才能切开)●减数分裂时的同源重组●启动重组的机制不同:双链断裂模型,但重组仅出现在断裂的一侧,杂合DNA只出现在同一条染色单体上●特异位点重组(site-specific recombination)●只发生在DNA的特异位点之间,依赖于序列的特异性,对序列的同源性要求低●λ噬菌体与大肠杆菌的位点特异性重组●特征:基本步骤同同源重组(链的交换、形成Holliday中间体、分支迁移、拆分),但是不需要RecA参与,分支迁移距离较短,依赖于特定的蛋白质识别重组位点,催化重组反应。
重组和转座

3. Rec A: ① RecA in bacteria has two quite different types of activity:
it can stimulate protease activity in the SOS response and can promote base pairing between a single strand of DNA and its complement in a duplex molecule.
基因转换
第三节 位点特异性重组
位点特异性重组最典型的列子是λ噬菌体对E.coli的整合。 1. att位点: 噬菌体λ的整合和切离是在特异位点即附着位点(attatchment sites, att ) 通过重组完成的,若这个位点发生突变就会失去功 能而抑制整合:
细菌 att B (BOB‟) 噬菌体 att P (POP’) “O” ,被称为核心(core)序列,为att B和att P共有的; 两侧序列B,B‟和P,P‟被称为臂(arms);
对靶位点的选择有三种形式:随机选择,热点选择
和特异位点的选择。
Insertion sequences have inverted terminal repeats and generate direct repeats of flanking DNA at the target site. In this example, the target is a 5 bp sequence. The ends of the IS consist of inverted repeats of 9 bp, where the numbers 1 through 9 indicate a sequence of base pairs.
第五讲 DNA的重组与转座

插入突变失活是转座的最直接效应,但转座 也可以通过干扰宿主基因与其调控元件之间的关
系或改变DNA的结构而影响基因的表达。
转座子插入靶序列后在受体DNA中形成3~12bp 的正向重复序列,在切离后可能就给DNA留下一小 段多余的靶位序列,从而导致编码的突变。
Meselson-Radding 模型
DNA单链形成可在一条DNA双链中首先产生, 随着缺刻处DNA链的修复合成,游离出的单链末端
侵入相邻DNA双链中,与同源部分配对结合,被替
换的DNA链形成一个逐渐增大的D-loop。D-loop断
开后产生的单链末端交叉侵入相邻DNA双链中,
DNA自由末端共价连接形成Holliday结构。
第五讲
DNA的同源重组与转座
基因重组概念
新的DNA分子中含有原来的两个DNA 分子 的片段。所有DNA均是重组DNA。 DNA分子内或分子间发重型遗传信息的重新 组合,称为遗传重组或基因重排。重组产物称
为重组DNA。
减数分
裂时,同源染色体在每一代都发生于有相同
RuvA RuvB
RuvC
特异位点重组
特异位点重组是指在重组酶介导下,在特异的重组位点间
发生重组,导致重组位点间交互交换的一种精确重组形式。
λ噬菌体的整合与切除
λ噬菌体编码λ整合酶(integrase),这个酶 能指导噬菌体DNA插入E.coli染色体中。
细菌的特异位点重组
如果一个DNA分子上两个特异位点之间发生重组,其后果 有两种可能性:两个位点之间的节段或被丢失,或被颠倒。有 些生物能够利用这种重组倒置来控制基因的表达。
真核生物的转座子
1.玉米的Ac-Ds调控系统
高中生物第十一节:基因重组与基因转位——基因重组概念
课次:22教学目的:使学生了解重组的四种类型,了解位点特异重组和异常重组的特点,掌握同源重组的机制。
重点:同源重组的机制难点:同源重组的机制复习旧课:提问1人,了解教学效果。
导入新课:第九章遗传重组第一节概述DNA分子内或分子间发生遗传信息的重新组合,称为遗传重组(genetic recombination)。
重组产物为重组体DNA (recombinant DNA) 。
DNA重组对生物进化起着关键的作用。
基因重组是指由于不同DNA链的断裂和连接而产生的DNA片段的交换和重新组合,形成新的DNA 分子的过程。
重组的类型:(1)同源重组(homologous recombination ):反应涉及到大片段同源DNA序列之间的交换。
其主要特点是需要RecA蛋白的介入。
(2)位点特异性重组(site-specific recombination):重组发生在特殊位点上,此位点含有短的同源序列,供重组蛋白识别。
(3)转座重组(transposition recombination):由转座因子产生的特殊的行为。
转座的机制依赖DNA 的交错剪切和复制,但不依赖于同源序列。
(4)异常重组分为两类,末端连接和链滑动。
其特征时重组对中很少或没有序列同源性,所以也称为非同源性重组。
第二节同源重组1 同源重组1.1 同源重组(homologous recombination):发生在同源DNA序列之间。
1.2 特征-进行同源重组的基本条件:1)在交换区具有相同或相似的序列:涉及同源序列间的联会配对,且交换的片段较大;单链DNA分子或单链DNA末端是交换发生的重要信号2)双链DNA分子之间互补碱基进行配对3)重组酶4)异源双链区的形成:涉及DNA分子在特定的交换位点发生断裂和错接的生化过程;存在重组热点。
e.g.:Euk.减数分裂时的染色单体之间的交换;细菌的转化,转导,接合,噬菌体重组同源重组是同源依赖性的,而非序列依赖性。
DNA的重组与转座培训

DNA的重组与转座培训DNA的重组与转座培训DNA的重组与转座是生物学中一项重要的技术,用于改变生物体的基因组,以实现特定的目的。
在这篇文章中,我们将详细介绍DNA的重组与转座技术,并提供相关实验操作的培训。
1. DNA的重组DNA的重组是指将来自不同来源的DNA片段进行重新组合,形成新的DNA序列。
这种技术的应用非常广泛,可以用于产生具有特定功能的蛋白质、疫苗的研发以及疾病的基因治疗等领域。
实验操作:步骤1:准备工作- 准备试管、酶切体系、反应缓冲液和DNA片段等。
- 温度、时间和酶切酶的浓度都是影响DNA重组效率的重要因素,所以需要根据具体实验来确定。
步骤2:酶切反应- 将待重组的DNA片段用限制性内切酶切剪,产生可粘性末端。
- 确保酶切反应的温度和时间符合要求。
步骤3:连接反应- 将两个经酶切的DNA片段混合,在酶切缓冲液中加入连接酶。
- 反应温度和时间的选择与步骤2类似。
步骤4:DNA重组产物的分离与提取- 通过琼脂糖凝胶电泳分离连接反应产物,并使用Gel Extraction Kit提取感兴趣的目标DNA。
2. 转座转座是指某些特殊的DNA序列(转座子)具有自主移动性,可以在基因组内发生位置的改变。
这种现象在进化中起到重要作用,并且可以被利用于基因突变的研究和修饰。
实验操作:步骤1:获得转座子DNA- 转座子可以在细菌、真核生物或植物中找到。
根据实验需要,可以使用PCR扩增或基因组DNA提取方法获得转座子DNA。
步骤2:构建转座载体- 选择合适的载体,将转座子DNA连接到载体上,并转化到适当的宿主细胞中,以扩增转座子。
步骤3:转座实验操作- 将转座载体引入到目标细胞中。
- 通过选择合适的培养基和筛选条件,筛选出带有转座子的目标细胞。
3. 实验操作的注意事项- 实验室操作要根据具体的实验方案和操作手册进行,严格按照操作流程进行。
- 培养容器、试剂和仪器要经过严格消毒,在实验操作时保持无菌环境。
分子生物学技术-第七章 DNA的重组与转座

指不含任何结构基因的转座子,它是一种自主 的单位,仅带有介导自身移动所需要蛋白质的基 因,是最简单的转座子。
IS 一般由一段DNA序列(一个转座酶基因,不 带其它标记基因)和两端反向重复序列组成,重 复序列一般在15-25bp。
不同IS插入位点不同,但序列大多为5-9bp, 转位发生后插入位点成为IS两端的同向重复序列。
一、霍利迪结构(Holliday Structure)
霍利迪结构:它是DNA重组过程中的一个中间体, 重组时两个DNA双链体以四股DNA在连结点形成的十字 形DNA分子构象。
异源双链 (Hetero duplex):指在重组部位,每条双 链中均有一段DNA链来自另一个双链中的相对链。
分支迁移(Branch migration):一旦两个重组的 DNA分子形成霍利迪结构,交叉连接点很容易像拉链一 样移动,从而扩大异源双链体区域,这一过程叫分支迁 移。
2、复合式转座子
它是一类较复杂的转座子,由两个重复序列 夹着一个或多个结构基因组成。除了含有转位相 关的功能外,还带有一些其他基因,如抗药性基 因、抗重金属离子基因等。
它们都具有末端反向重复序列、可编码基因 和插入位点的正向重复序列
TnA转座子:
结构较复杂,长度约5000bp。两端没 有IS结构,仅含有两个末端反向重复序列, 一般约38bp。内部通常含有转位相关的基 因和抗药性基因等,插入位点约5bp。
第七章 DNA的重组与转座
重组分为4类:
同源重组:重组的DNA之间具有同源性 位点特异性重组:重组DNA之间不需要同源,但
是需要识别特异的DNA序列 转座重组:需要转座子
异常重组:不正常细胞加工的结果。
2-染色体与DNA-DNA修复与转座

2 DNA的损伤修复
由于染色体DNA在生命过程中占有至高无上的地位, DNA复制的准确性以及DNA日常保养中的损伤修复就有 着特别重要的意义。
大肠杆菌中DNA的修复系统
2.2 DNA损伤修复的类型
2.2.1 2.2.2 2.2.3 2.2.4 2.2.5 2.2.6 错配修复 碱基切除修复 核苷酸切除修复 DNA直接修复 重组修复 SOS反应诱导的修复
该系统识别母链的依据来自Dam甲基化酶,它能使位于 5’GATC序列中腺苷酸的N6位甲基化。一旦复制叉通过复制起 始位点,母链就会在开始DNA合成前的几秒种至几分钟内被甲 基化。
只要两条DNA链上碱基配对出问题,错配修复系统就会根据 “保存母链,修正子链”的原则,找出错误碱基所在的DNA链, 并在对应于母链甲基化腺苷酸上游鸟苷酸的5’位置切开子链,再 启动特定的修复途径,合成新的子链片段。
2.6 DNA的转座 (DNA Transposition)
DNA的转座,或称移位(transposition),是由可 移位因子(transposable element)介导的遗传物 质重排现象。
转座过程示意图
2.6.1 转座子的分类和结构特征 (transposon,Tn)
转座子: 是存在于染色体DNA上可自主复制和位移的基本单 位。
DNA的重组修复
2.2.6 SOS反应诱导的修复 SOS response & DNA repair
• SOS反应:是细胞DNA受到损伤或复制系统受到抑制的紧急情况 下,为求得生存而出现的应急效应。 • SOS反应诱导的修复系统包括避免差错修复(error free repair)和 易产生差错修复(error-prone repair)
2.6.2 转座作用的机制
基因组重组和转座子活动

基因组重组和转座子活动作为生命的基本单元,细胞在生长发育、适应环境等过程中需要不断地调控基因表达。
而基因组重组和转座子活动作为基因表达中的一个重要过程,也在细胞发育和进化过程中发挥着重要作用。
一、基因组重组基因组重组是指在细胞分裂过程中,染色体上一段DNA序列与另一段DNA序列交换位置的现象。
这一过程可以发生在减数分裂过程中,也可以发生在有丝分裂过程中。
在减数分裂过程中,有趣的是,同源染色体之间的互换过程可以促进基因的多样性,对在进化过程中的适应性有重要作用。
同样,在有丝分裂过程中,由于在细胞周期的S期复制中,丝粘连复合物的结构可以导致同源染色体之间的交叉而重组,从而形成新的基因组序列。
基因组重组既重要又复杂,因此,它需要细胞中的一些特殊酶来协助完成,包括重组酶、拼接酶和核酸酶等。
二、转座子活动转座子是指在基因组中具有自主转移能力和引起基因组重组的DNA片段。
插入到一个基因区域可以革命性地改变基因的表达,所以转座子的活动可以起着丰富、多样化物种基因组的作用。
然而,由于转座子的位置特异性和非特异性种族特异性的现象时有喜见,转座子也可以带来某些负面效应,如遗传疾病和基因突变。
转座子可以通过不同的机制来使它们移动和影响基因组。
其中,一种被称为复制粘贴转移机制。
这种机制涉及在高危核酸中逆转录酶的作用下,从转座子中复制DNA片段,并插入到宿主的相同或不同部位。
另一种机制被称为切-粘式转移机制,其中转座子内部的酶可以切除DNA,然后插入到宿主某个区域的特定位点。
三、基因组重组和转座子活动协同作用基因组重组和转座子活动是不可分割的。
转座子的运动和基因组的重新组合是一个复杂的动态过程,对于各种生命现象都有很大的影响,包括不同物种之间的进化、种群的遗传多样性、染色体数目的变化、疾病的产生等。
例如,转座子可以插入到调控基因的区域,导致这些基因的表达被调节。
基因组重组可以导致整个基因和调控区域之间的距离发生变化,从而影响基因组的整体表达。
第七章 DNA重组与转座

转座作用的遗传学效应
① 转座引起插入突变 ② 转座产生新的基因 ③ 转座产生的染色体畸变 ④ 转座引起的生物进化.
转座子的转位特征 P204
7.4 逆转录病毒和逆转录转座子
以RNA介导的转座与逆转录病毒有关,被转移 的因子称为逆转录转座子。
逆转录转座子是真核生物转座子的重要类型。
逆转录病毒与逆转录转座子的区别:
来源于宿主dna普遍性转导generalizedtransduction局限性转导restrictedtransduction完全转导外源性dna片段与受体菌的染色体整合并随染色体而传代称完全转导流产转导外源性dna片段游离在胞质中既不能与受体菌染色体整合也不能自身复制称为流产转导局限性转导或特异性转导所转导的只限于供体菌染色体上特定的基因
局限性转导或特异性转导, 所转导的只限于供体
菌染色体上特定的基因。如λ 噬菌体进入大肠埃希
菌。
4.细菌的细胞学机制
P193
7.2 位点专一性重组:导致DNA序列发生重排,不依赖于DNA 顺序的同源性,而依赖于能与某些酶相结合的特异DNA序列, 这些酶能催化DNA链的断裂和重接,引发位点专一性重组。
融合在一起,转录后形成一条即含病毒序列又含
C-onc的RNA,后者如含包装信号可被包装进
病毒颗粒中,形成含有一条融合RNA和一条正
常病毒RNA的二倍体病毒颗粒。 二者分子之间发生重组将会产生转化逆转录 病毒基因组。病毒携带V-onc,即具有转化能力。
逆转录病毒是具有感染能力的病毒颗粒,可以 在细胞之间转移。 逆转录转座子是宿主DNA基因组的组分,可以 在基因组内转座,但不能在细胞之间转移。
a)DNA转座子 DNA DNA 细菌转座子及插入序 列
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
部位被切断,然后暴露的末端侵入到另一个双链
DNA分子,发现它的互补序列以后将替代与它相
同的链。然后DNA聚合酶将用留下的那条链作为
模板,填充上侵入链所留下的缺口。被替代的链 被降解,它的残留末端与另一分子中新合成的 DNA链相连接形成Holliday连接体。
27
28
30
• Holliday连接体形成以后,就会产生异构化然 后拆分。 • 单链侵入模型中,异源双链首先只在两个 DNA分子中的一个形成。Holliday中间体形成 以后通过支链迁移能够在另一个DNA分子上 产生异源双链,这样就解释了异源双链是如何 形成的。
• 同源性重组最主要特征是相关的酶可以利用任 何一对同源序列为底物。
10
• 真核生物减数分裂时的染色体之间的交换, 某些低等真核生物及细菌的转化、转导、 接合,噬菌体的整合等都属于同源性重组 这一类型。 • 在整个基因组中,同源重组的频率并不恒 定,并且跟染色体的结构有关。例如在异 染色体附近遗传物质的交换要受到抑制。
50
51
53
52
54
53
55
第二节 位点特异性重组
• 位点特异性重组是指发生在DNA特异性位点上的
重组。
• 与同源重组不同,位点专一性重组不是由DNA序
列的同源性所引导,而是在结合序列部位由专一
的酶来催化断裂重接。位点专一性重组是由使相 应酶出现的调节过程所引发,而同源重组则是由 能与RecA蛋白结合的DNA序列随机断裂引发的。
• 通过同源性重组将DNA序列引入真核基因组已 经为研究高等生物开辟了一条新途径。
8
• 用人工操作构建DNA重组体,将其导入受
体细胞扩增或表达,称为DNA克隆或分子
克隆(cloning)。
9
第一节 同源重组
• 同源重组(homologous recombination)发生 在两个同源DNA片段之间,是在两个DNA分 子的同源序列之间直接进行交换的一种重组形 式。同源重组主要是利用DNA序列的同源性识 别重组对象,由碱基序列提供识别的特异性。
24
25
27
• Holliday认为在DNA分子上存在某些位点,特 殊的引发重组的酶能够识别这些位点,确保两 条链在相同的部位被切断。
• 目前还没有足够的证据证明这些位点的存在。
26
(2)单链侵入模型
• 单链侵入模型是对Holliday模型的修改。
• 在该模型中,两个DNA分子中的一个单链在随机
双螺.旋中的同源序列发生置换,这一反应被称为 单链摄取(single-strand uptake)或单链同化(singlestrand assimilation)。
44
45
46
47
3. Ruv蛋白和 Holliday连接体的拆分
• 大肠杆菌有一组由三个基因编码与随后的重组相 关联的蛋白。
• ruvA 和 ruvB 基因的产物促进异源双链的形成。
34
35
37
36
38
大肠杆菌重组的分子基础
• 在分子水平上,重组事件发生的先后顺序是相似
的:从一个已断裂的分子中产生的单链与其对应 的双螺旋发生作用,配对区间扩展,重组中间体 形成直到核酸内切酶解离此两个双螺旋分子。 • 识别反应是重组机制的不可分割的重要部分,并 且只涉及到DNA分子的特定区域而不是整个完整 的染色体。
Ruv A 蛋白可以识别Holliday连接体的结构,Ruv
B是一个腺苷三磷酸酶并可能提供支链迁移的动 力。
48
• Ruv A在交叉点处与DNA所有的4条链相结合,在
交叉点的上游,两个Ruv B六聚体环状结构分别 与每个双螺旋相结合。
• RuvAB蛋白复合体可以使支链以10~20 bp的速度
迁移。
11
1. 进行同源重组的基本条件
(1)在交换区具有相同或相似的序列
• 发生同源性重组的两个区域的核苷酸序列必须 是相同或非常相似的。
• 同源性重组常常只发生在两个DNA分子中的相 同部位。
12
(2)双链DNA分子之间互补碱基进行配对 • 两个DNA分子的链之间互补的碱基配对确保重
组只发生在同样的基因座之间。
• RecA可以促进单链DNA 与其在双链DNA 分子中 互补的DNA 链的碱基相配对。 • RecA还可以在SOS 反应中激活蛋白酶。 RecA 需 要单链DNA和 ATP才能激活蛋白酶。
43
• RecA能够利用由RecBCD在Chi位点附近切割所释
放的单链3’末端。 • RecA的DNA操作酶活性可以使一个单链DNA与
31
32
34
33
35
• D环在DNA聚合酶的作用下修补而扩展,直至D
环的长度与“受体”DNA双螺旋缺口长度相当,
互补的两条单链退火。此时在缺口的两侧各有一
段异源双链DNA,两个杂合双螺旋之间为断裂 的缺口,并且此缺口为供体DNA序列所填充。 • 缺口处双链的完整性可以被以缺口3’端为起始的 修补合成来修复。缺口是被两次单链DNA的合 成而修复的。
57
λ噬菌体DNA的整合与切除
• 为了进入溶源状态,游离的 DNA必须整合 (intergrate)到细菌DNA中去;而为了从溶原状 态向裂解状态转化,原噬菌体DNA则必须从细菌染 色体DNA上切除(excise)。
• 整合和切除均通过细菌DNA 和λDNA上特定位点—
附着点(attachment site,att)之间的重组而实现。
29
⑤
⑦
30
(3)双链断裂修复模型
• 目前证明通过两个DNA分子之中的一个双链断裂引
发重组是个常见的机制。
• 该模型认为参与重组的两个DNA分子之一的两条链
被核酸内切酶切断,然后在核酸外切酶作用下产生3’
单链黏性末端。
• 两个3’游离末端之一侵入到另一个双螺旋的同源区,
置换“供体”双螺旋的一个单链而形成一段异源双 链DNA,并同时产生一个D环(D-loop)。
49
• 第三个基因ruv C编码一种能够特异性地识别 Holliday连接体的核酸内切酶,此酶可以在体外切 断此种交叉而拆分重组中间体。 • 一个含4个碱基的序列提供了Ruv C拆分Holliday连 接体的热点。这一序列决定了拆分过程中哪一对 DNA链被切断,从而决定发生的是补丁型重组 (patch recombination)还是剪接型重组(splice recombination)。
• 分子间位点专一性重组产生整合或融合; • 重组位点的正向顺序间的分子内位点专一 性重组产生剪切和解离; • 重组位点的反向顺序间的分子间位点专一 性重组产生倒位。
56
位点特异性重组的功能:
1. 调节噬菌体DNA与宿主菌染色体DNA的
整合; 2. 调节特定基因的表达;
3. 调节胚胎发育期间程序性的DNA重排。
recombination)
• 异常重组(illegitimate recombination)
• 转座重组(transposition recombination)
7
• 许多重组反应利用不止一种机制进行,例如免
疫球蛋白VDJ连接涉及到位点特异性识别和类
似异常重组的过程;酵母接合型的转换既有同
源性重组和位点特异性重组的特征,也具有复 制转座过程的特征。
54
• 位点特异性重组最早是在λ噬菌体的遗传学研究中
发现的。
• λ噬菌体DNA通过重组整合到大肠杆菌染色体的
专一性位点上,转变成为原噬菌体DNA的非活性
状态。
• λ噬菌体有两种存在型式:裂解状态和溶源状态。
两种类型间的转换是通过位点特异性重组实现的。
55
根据重组所涉及的体系和重组位点的方向,
位点专一性重组可以介导3类反应:
单链的断裂,从而引发重组。
• 两个断裂单链的游离端彼此交换,形成两个异源
双链,然后末端连接形成Holliday连接体
(Holliday Junction)。
17
18
20
• 一旦Holliday连接体形成后,它能进行重排从 而改变链的彼此关系。这种重排称为异构化, 因为在此过程中没有键的割裂。 • 一旦形成Holliday连接体后,就能被拆分。是
减数分裂和体细胞核基因中发生,也在线粒
体基因间和叶绿体基因间发生。
5
• DNA重组的意义是能迅速增加群体的遗传 多样性,通过优化组合积累有利突变,推 动生物进化。此外,DNA重组还参与DNA 损伤修复,某些基因表达的调控等重要的 生物学过程。
6
根据不同的机制,可将重组分成4类: • 同源性重组(homologous recombination) • 位点特异性重组(site-specific
37
1. chi位点和RecBCD核酸酶
• 在λ噬菌体的一些突变种内,存在一些称为 chi的位点。chi位点单一的碱基对的改变即 可激发重组的发生。这些位点皆含有一个
恒定的非对称的8bp序列:
5’ GCTGGTGC 3’ 3’ CGACCACC 5’
38
• Chi位点是一个由基因recBCD编码的Rec BCD酶 作用的靶部位。
• RecBCD酶是一种多功能酶,具有几种不同的活
性。它是一个强有力的降解DNA的核酸酶,早期
被检定为活性核酸外切酶V,具有解旋酶的活性。
• RecBCD 所介导的解旋和切割可以被用来产生末 端,并引发异源双链的形成。
39
40
42
41
43
42
44
2. RecA和Holliday连接体的形成
RecA具有两种相当不同的活性:
• 在该部位两个双链DNA分子通过链之间互补的 碱基配对被维系在一起称为联会。
13
(3)重组酶