2014 海产品中的砷及其代谢机制的研究进展_王瑛
“海藻中砷.铝、镉形态分析技术研发与应用”成果通过专家鉴定

在影 响研 究与评价 ”等2 项 自然科 学基金项 目完成情 况的汇报 ,审 阅了相关验收材料 ,通过质询和讨论 ,认为这4 个项 目均顺利完成 了合 同规定的各项指标 ,且 项 目管理规范 ,经费使用合理 技术研发与应用”成果通过专家鉴定
质信息 采集系统 、水质智 能预警系统 、分布 式集 成控制系统等 关键技术难点 ,开展 以数字化技术对集 约化养殖所涉 及
的对象和 全过程进行数字化 和可视化表达 、设计 、控制 、管理 的研 究与应用 。研 究成果包括智能化 投饲 、增氧调控 、 远程监控 等智能化控制 系统装备 ,并大面积 应用于池塘 、工厂化和网箱养殖 ,推广 示范应用范 围包 括上海 、广东 、北 京 、新疆 、河南 、福建 、宁夏 、浙江 等地 。经实际生产证 明,池塘养殖节 约用电超 过4 0 %,节省人力成本5 0 %以上 ,经 济效益增加 1 0 %,技术稳定可靠 ,管理维护成本低 ,具有 良好的社 会效益和应用前景 。(
通过 “ 提 高斑节对 虾精卵 质量 的研 究 ”项 目实 施 ,揭示 了人 工养殖条 件下斑 节对虾 生长和 发育及 性成熟 特点 、
亲虾性腺 发育 和关键营养物质 的关系 、生化组成 及内分泌激素 的变 化特征 ,形成 了斑 节对 虾亲虾全人工培 育的基础理
浙江省沿海常见海产品总砷含量水平调查分析
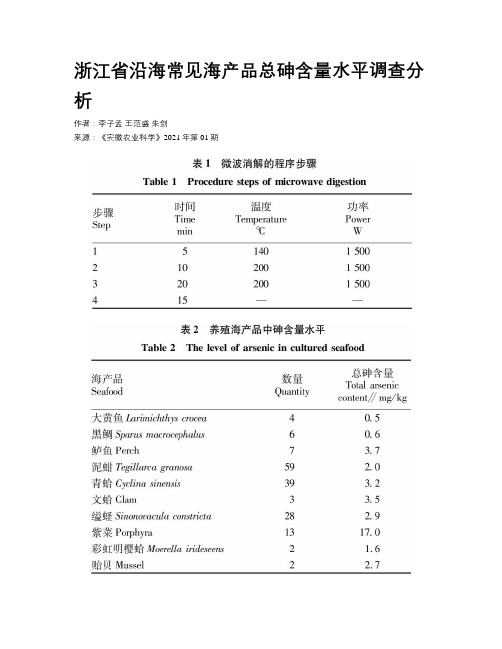
浙江省沿海常见海产品总砷含量水平调查分析作者:李子孟王范盛朱剑来源:《安徽农业科学》2021年第01期摘要采用电感耦合等离子体质谱法(ICP-MS)对浙江省沿海常见海产品中的总砷和无机砷含量进行测定,采集并检测27种775个海产品样品,结果显示,不同种类海产品的总砷含量均值在0.5~17.0mg/kg,其中总砷含量最高的是紫菜,均值达17.0mg/kg,其次为甲壳类、头足类和贝类,鱼类总砷含量最低。
根据不同种类海产品砷迁移规律和生活习性,推断藻类中总砷含量水平主要与它本身的细胞壁特性相关;贝类中总砷含量对沉积物中砷的富集率为27%~58%,蛤蜊相比其他贝类对砷的富集能力更强,为文蛤、青蛤总砷含量较高的主要原因;甲壳类中日本鲟和中华管鞭虾相对砷含量较高是因为較广的食性范围;鲈鱼为凶猛肉食性鱼类,总砷含量高的主要原因是摄食富集。
关键词总砷;ICP-MS;含量测定;海产品;浙江省沿海中图分类号TS254.7文献标识码A文章编号0517-6611(2021)01-0186-02doi:10.3969/j.issn.0517-6611.2021.01.050AbstractIn this study, the content of total arsenic and inorganic arsenic in the common marine products of Zhejiang Province was determined by inductively coupled plasma mass spectrometry (ICP-MS). 27 kinds of 775 samples of marine products were collected and detected. The results showed that the average content of total arsenic in different kinds of marine products was between 0.5-17.0mg/kg, among which Porphyra was the highest with the average content of 17.0mg/kg,followed by crustaceans, cephalopods and shellfish, and the content of total arsenic in fish was the lowest among the three species. According to the migration rule and living habit of arsenic in different kinds of seafood, it was inferred that the level of total arsenic in algae was mainly related to its own cell wall characteristics;the enrichment rate of total arsenic in shellfish to arsenic in sediment was 27%-58%, and the enrichment ability of clam to arsenic was stronger than other shellfish, which was the main reason for the high content of total arsenic in clam and Cyclina sinensis;the relatively high arsenic content of Japanese sturgeon and Chinese tube prawns in crustaceans was due to the wide range of food habits;Perch was a fierce carnivorous fish, the main reason for the high total arsenic content was the enrichment of food.Key wordsTotal arsenic;ICP-MS;Content determination;Seafood;Zhejiang coastal area目前,居民在日常饮食生活中的主要消费海产品包括鱼、虾、蟹、贝4类。
微生物砷还原机制的研究进展

微生物砷还原机制的研究进展陈倩;苏建强;叶军;朱永官【摘要】砷是一种剧毒物质,环境中的砷对人体健康存在潜在威胁,因此长期以来备受关注.微生物的各种代谢过程对砷在环境中的归趋起着重要作用,其中砷还原微生物能将吸附于固体矿物中的As(Ⅴ)还原为可溶性强的As(Ⅲ),使砷进入液相,从而加剧了地下水等饮用水源的砷污染.论文主要介绍了两种微生物砷还原机制(异化砷还原和细胞质砷还原)在作用机理、蛋白质结构和表达调控等方面的研究进展,旨在为更深入理解微生物介导砷的生物地球化学循环以及砷污染的微生物修复技术提供参考.【期刊名称】《生态毒理学报》【年(卷),期】2011(006)003【总页数】9页(P225-233)【关键词】砷还原;微生物;异化砷还原;细胞质砷还原【作者】陈倩;苏建强;叶军;朱永官【作者单位】中国科学院城市环境研究所中国科学院城市环境与健康重点实验室厦门361021;中国科学院城市环境研究所中国科学院城市环境与健康重点实验室厦门361021;中国科学院城市环境研究所中国科学院城市环境与健康重点实验室厦门361021;中国科学院城市环境研究所中国科学院城市环境与健康重点实验室厦门361021【正文语种】中文【中图分类】X171.5砷是自然界中普遍存在的有毒类金属。
1993年世界卫生组织将饮用水中的砷标准由原来的50 μg·L-1降为10μg·L-1。
2006年美国环保局将其列为第一位饮用水污染物[1-2]。
我国从 2007年 7月开始也以10μg·L-1作为生活饮用水中允许的砷浓度上限。
砷在环境中共有 4种价态:As(-III)、As(0)、As(III)和As(V),其中以As(III)和 As(V)较为常见,它们也是造成环境砷污染的主要形态[3]。
微生物在长期的历史演化过程中进化出了各种抗砷机制,更有最新的研究表明有细菌 (Halomonadaceae GFAJ-1 strain)能以砷代替磷作为细胞中DNA的组成成分供细菌生长[4],但是这项研究发表后亦颇有争议,需要有进一步可靠的数据支持。
砷在水生生物中的生物累积、转化及在其他生物体内的代谢毒理学研究进展

DOI:10.7524/j.issn.0254-6108.2022100401冉茂霞, 吴迪, 史永富, 等. 砷在水生生物中的生物累积、转化及在其他生物体内的代谢毒理学研究进展[J]. 环境化学, 2024, 43(4): 1069-1084.RAN Maoxia, WU Di, SHI Yongfu, et al. Research progress on arsenic's bioaccumulation and biotransformation in aquatic organisms, and its metabolism and toxicology in other organisms[J]. Environmental Chemistry, 2024, 43 (4): 1069-1084.砷在水生生物中的生物累积、转化及在其他生物体内的代谢毒理学研究进展 *冉茂霞1,2 吴 迪1,2 史永富1 ** 顾润润1 黄冬梅1 席寅峰1 汤云瑜1 叶洪丽1 张 璇1 方长玲1(1. 中国水产科学研究院东海水产研究所,农业农村部远洋与极地渔业创新重点实验室,上海,200090;2. 上海海洋大学食品学院,上海,201306)摘 要 砷作为全球水生生态系统的重要污染物,普遍存在于淡水和海洋环境中,具有一定的生物累积性与生物毒性. 水生生物作为生态系统的重要组成部分,砷在全球生物化学循环的过程中通过迁移、累积、转化、富集在水生生物体内,产生毒性作用,而砷对水生生物的毒性与其在水生生物中的赋存形态有关. 目前关于砷在水生生物中的生物累积、生物转化及其代谢毒理的基础科学研究受到国内外研究者的广泛关注,而少有对该领域内的研究进展、研究热点、趋势方向的系统整合,本文针对这一领域文献进行梳理,系统阐述了砷在水生生物中的生物累积和生物转化情况;概述了砷在水生生物、小鼠/大鼠、人体、微生物等生物中的毒性作用及代谢机制;并提出了未来有关砷研究可关注的重点及方向. 本文可为进一步阐明砷在水生生物中的生物累积转化规律及砷的代谢毒性作用,为研究者进一步深入探索该领域内科学问题提供参考资料,同时对水产品安全、环境生态、医学等相关领域的研究具有一定的借鉴意义.关键词 砷,水生生物,生物累积,生物转化,毒理效应,代谢机制.Research progress on arsenic's bioaccumulation and biotransformation in aquatic organisms, and its metabolism and toxicologyin other organismsRAN Maoxia1,2 WU Di1,2 SHI Yongfu1 ** GU Runrun1 HUANG Dongmei1 XI Yinfeng1 TANG Yunyu1 YE Hongli1 ZHANG Xuan1 FANG Changling1(1. Key Laboratory of Oceanic and Polar Fisheries, Ministry of Agriculture and Rural Affairs, China, East China Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Shanghai, 200090, China;2. College of Food Science andTechnology, Shanghai Ocean University, Shanghai, 201306, China)Abstract Arsenic is a bioaccumulative and biotoxic pollutant in global aquatic ecosystems and can be ubiquitously found in freshwater and marine environments. In the global biochemical cycle process, Arsenic can migrate, bioaccumulate, transform, and accumulate in aquatic organisms, resulting in toxic effects. The toxicity of Arsenic is related to its valences. Various studies have focused on preliminary research of Arsenic's behaviors and metabolic toxicology in aquatic2022 年 10 月 4 日 收稿(Received:October 4,2022).* 中国水产科学研究院东海水产研究所基本科研业务费资助(2021M05).Supported by the Fundamental Research Funds of the East China Sea Fisheries Research Institute of China (2021M05).* * 通信联系人 Corresponding author,E-mail:*****************1070环 境 化 学43 卷environments. Still, a systematic review is not yet performed on the research progress, highlights, and trend. This paper summarized published reports and detailed the bioaccumulation and biotransformation processes of Arsenic in aquatic environments. It also addressed its toxic effects and metabolic mechanisms in aquatic biota, mice/rat, human, microorganisms, and other organisms. Also, we proposed the emphasis and direction of future work on Arsenic. This paper could serve as reference material for researchers exploring scientific issues in this and other close research fields, such as aquatic product safety, environmental ecology, and medicine.Keywords arsenic,aquatic organisms,bioaccumulation,biotransformation,toxic effects,metabolic mechanisms.砷是全球水生生态系统的重要污染物,是一种能通过地质过程或人类活动在水生环境中释放的有毒元素,一些人类活动如过矿石冶炼、煤炭燃烧加剧了自然环境中的砷浓度[1 − 3]. 环境中的砷可分为无机砷和有机砷,无机砷主要包括砷酸盐(As(Ⅴ))、亚砷酸盐(As(Ⅲ)),有机砷主要包括砷甜菜碱(AsB)、砷胆碱(AsC)、甲基砷酸(MMA)及其甲基化砷复合物(MMAⅢ、MMAⅤ)、二甲基砷酸(DMA)及其二甲基化砷复合物(DMAⅢ、DMAⅤ)、砷糖(AsS)和砷脂(AsL)[4]. 砷在水生生物中的赋存形态(图1)因不同的水生生物而有所不同,水生生物普遍对砷具有较高的累积能力,可通过摄食、皮肤渗透或者黏膜吸收砷,并能通过主动运输进入细胞[5],吸收的砷会对机体造成不良影响,如中毒、免疫紊乱、组织损伤以及细胞死亡[6],而其毒性大小又与存在形态有关,由半数致死剂量(LD50)确定的不同形态的砷毒性顺序为As(Ⅲ)>As(Ⅴ)>MMA V>DMA V>AsS>AsC>AsB[7 − 8],通常而言,无机砷毒性高于有机砷毒性,As(Ⅲ)毒性高于As(Ⅴ),但在比较各种砷化合物对人肝细胞毒性实验时发现,毒性顺序为DMAⅢ≫As(Ⅲ)>MMAⅢ>As(Ⅴ)>MMAⅤ=DMAⅤ[9 − 10],因此,砷对生物体的毒性作用引起了全球范围内的关注.图 1 典型砷化合物的化学结构式Fig.1 Chemical structures of typical arsenic compounds无机砷可在水生环境中经水生生物及微生物代谢转化形成多种形态的砷,这些形态的砷毒性小于原有的无机砷毒性[11],说明砷在形态上的转化可能是砷的一个重要解毒机制,因此,关于砷毒性及其代谢转化的关系一直是重要的研究领域. 孟加拉国、印度、美国、阿根廷、智利、中国等都遭到了由砷污染造成的健康问题[12],作为一种自然污染物,砷已被证明对人类健康造成不利影响,其中无机砷可导致皮肤癌、肺癌、膀胱癌、神经退行性等疾病[13],已被国际癌症研究中心(IARC)列为Ⅰ类致癌物[14]. 有研究表明,砷暴露与神经退行性等疾病发病率增加有关,然而其毒性致病机理尚不清楚,因此国内外大量研究者采用动物模型研究神经退行性等疾病(如阿尔兹海默症、帕金森等)或缓解此类疾病的药物,同时,近年来有许多砷与微生物的研究值得关注. 在医学研究中发现,肠道微生物与砷代谢有着极大的相关性;在环境学方面,砷的微生物参与了全球砷的生物化学循环过程,能吸收转化环境中的砷,一定程度上降低了环境中的砷污染,具有极大的生物治理潜力[15 − 17]. 因此,近年来,有大量关于砷在小鼠/大鼠、人体、微生物中的代谢毒理情况的研究,可为人类流行病学、毒理学和生态环境研究奠定坚实的4 期冉茂霞等:砷在水生生物中的生物累积、转化及在其他生物体内的代谢毒理学研究进展1071基础.全球水生生态系统中的砷在不同水生生物中的生物累积、生物转化、产生的毒性及毒理效应的过程是复杂的,不同的水生生物对砷的耐受程度不同,不同形态的砷对水生生物产生的毒性也不同,因此,砷在水生生物中的生物累积、生物转化及毒理效应受到国内外研究者持续而广泛的关注,大量实验研究集中砷在水生生物、小鼠/大鼠、人体、微生物中的代谢毒理学,本文将重点对这一系列热点问题展开综述,可为进一步阐明砷在不同生物中的累积转化过程及毒理学效应,为研究者深入探索该领域提供一定的参考.1砷在水生生物中的生物累积和生物转化(The bioaccumulation and biotransformation of arsenic in aquatic organisms)1.1 砷在水生生物中的生物累积1.1.1 砷在鱼类中的累积通常来说,鱼类处于水生食物链的顶端,在正常的新陈代谢中会从食物、水、底泥环境中不可避免地累积重金属元素,因此,鱼类常被用做极具代表性的生物标志物,从而评价鱼类生活环境的金属水平. 淡水生物和海水生物的砷形态形成有所不同,鱼和贝类中砷形态和含量随着个体的种类和采样面积存在显著差异,砷甜菜碱(AsB)是鱼类中的主要砷形态[18].无机砷对水生生物有剧毒,但膳食无机砷暴露对鱼类的慢性影响尚不清楚. 为了解慢性砷暴露下鱼类中砷的累积情况,Chen等[19]对淡水青鳉(Oryzias mekongensis)进行一项为期28 d的慢性无机砷膳食暴露(As(Ⅲ)、As(Ⅴ)),发现经同浓度(1.3 mmol·L−1)的As(Ⅲ)、As(Ⅴ)分别暴露后,淡水青鳉组织中砷的生物累积浓度先增后减,而这两种浓度先增后减的时间存在一定的差异性. Di等[20 − 21]在对鲫鱼(Carassius auratus)进行一项为期40 d的慢性无机砷(As(Ⅲ)、As(Ⅴ))食物相暴露研究中也发现了类似的相关性,在分别暴露相同浓度(50 μg·g−1)的As(Ⅲ)、As(Ⅴ)后,测定鲫鱼肝脏及全身肌肉中As(Ⅲ)、As(Ⅴ)和AsB的累积情况,在鲫鱼肝脏中,AsB在暴露过程中比例呈上升趋势(34%—66%),As(Ⅲ)、As(Ⅴ)比例在暴露过程中逐渐下降(84%—91%). 在对鲫鱼肌肉组织中砷累积情况的研究发现,经As(Ⅲ)暴露的鲫鱼肌肉中检测到更高比例的AsB,两种无机砷累积的减少与其转化为无毒的AsB的增加有关,相比于As(Ⅴ),As(Ⅲ)更容易转化为AsB. 在暴露的食物相中,As(Ⅲ)的生物利用度低于As(Ⅴ),这可能是由于As(Ⅲ)通过鲫鱼肌肉组织的能力较低. 而在这项研究[21]中发现, 鲫鱼肌肉组织含有大量的As(Ⅲ),说明As(Ⅴ)能大量转化为As(Ⅲ),从而累积在鲫鱼肌肉组织中.另外,砷在鱼类各组织中的累积情况不同,且不同品种间累积情况存在差异. Kim等[22]对幼年岩鱼进行水环境砷暴露20 d,发现在幼岩鱼组织中砷的累积顺序大小为肝>肾>脾>鳃>肠>肌肉. Kumar等[23]采用亚砷酸钠对鲶鱼(Clarias batrachus)进行为期60 d的暴露,最后发现砷在鲶鱼各组织中的累积情况为肝脏>血液>肌肉>皮肤>大脑. Juncos等[24]发现,克里奥尔鲈鱼、虹鳟鱼、褐鳟鱼不同组织累积砷浓度大为肾脏>肝脏>鳃>肌肉. 作为高等物种水生食物的重要来源,水环境中的无脊椎动物易富集砷[25],鱼类存在于水生食物链的顶端,在捕食这些无脊椎动物时同时摄入砷[26]. 大量的研究表明,AsB是这些鱼类组织中砷的主要存在形态,而无机砷(As(Ⅲ)、As(Ⅴ))和有机砷(MMA、DMA、AsC)占总砷的比例相对较少. 这些砷形态在鱼体内累积、储存、进一步转化,通过食物链传递,产生生物放大作用,累积至人体中,对生物与人类健康产生潜在危害[27 − 28],因此,鱼类是人类砷暴露的重要来源.1.1.2 砷在甲壳类、双壳类水生动物中的累积甲壳类、双壳类水生动物由于自身滤食性的特点,易富集重金属元素,AsB是最主要的砷形态[29].例如,Zhang等[30]在对中国南方湛江河口地区的甲壳类动物中的砷含量测定发现,AsB占所有砷化物的80.6%—98.8%. Hong等[29]对韩国浦项市高度工业化地区中水生生物中的砷累积研究中发现,在蟹类和虾类中AsB占总砷浓度的88%,相比之下,甲壳类、双壳类水生动物中累积的砷浓度高于鱼类. 另外,甲壳类水生动物不同组织对砷的累积情况有所不同,Devesa等[31]测定克氏原螯虾整个生物体及其各个部分的总砷和砷形态的浓度,发现无机砷占总砷的18%—34%,不同部位的砷浓度与克氏原螯虾的部位有关,其中肝胰腺的总砷及无机砷含量最高,尾巴中的总砷和无机砷含量最低. 同样地,Liao等[32]将南美白对虾分别暴露于不同浓度的As(Ⅲ)中21 d,发现肝胰腺也是南美白对虾体内累积砷的主要组织,总砷和无机砷的累积浓度同暴露浓度呈正相关. 而累积高砷浓度的肝胰腺会对甲壳类动物的繁殖产生影响,这与Yamaguchi等[33]发现砷对野生淡水蟹的性腺发育有影响的研究结果一致.人群通过膳食摄入鱼类、甲壳类、双壳类及其制品,可使砷在消费者体内蓄积暴露,而甲壳类、双壳类水生动物则经代谢转化形成大量的AsB,以其作为机体内主要的砷形态,使机体免受毒性更高的无机砷的损害作用,这可能是水生生物的一种自我保护机制. 同时,对于甲壳类水生动物的研究发现,不同种类和不同组织中砷的累积情况均不同,但大部分甲壳类水生动物的肝胰腺中都累积了大量的砷,这或许是因为肝胰腺作为甲壳动物最重要的组成部分,具有免疫砷毒性及解毒作用.1.1.3 砷在藻类中的累积PO3−4PO3−4PO3−4PO3−4除鱼类、甲壳类、双壳类水生动物易累积环境中的砷外,水生环境中的藻类同样暴露于重金属污染中,易累积砷,研究发现大型藻类对砷具有较高的累积速率和对金属的亲和力,可以高度暴露于砷的污染中[34]. 但影响海藻中砷累积的因素有很多,例如,藻类的种类、环境中的砷浓度、藻类的收获时间、藻类对养分的吸收程度和环境中的温度状况. 虽然海藻中的有机砷含量高于无机砷,但由于海藻中的无机砷比有机砷毒性更大,因此研究海藻中的无机砷比有机砷更为广泛[7]. 对海藻累积砷的研究过程中发现,海藻酸盐是褐藻活细胞中最丰富的多糖,能吸收环境中的重金属和类金属[35],导致褐藻中累积砷. 而外界磷酸盐()的浓度也可能促进海藻对砷的吸收,因为磷酸盐()和砷酸盐As(Ⅴ)结构相似,它们具有几乎相同的p K a值,相似的带电氧原子以及相似的热化学半径,因而藻类不能区分这两者[36 − 38],并且随着环境中浓度的增加,微藻对砷的吸收会受到抑制[39],这可能是由于浓度的增加会减少细胞膜上As(Ⅴ)转运体的数量或者是其与As(Ⅴ)竞争细胞质As(Ⅴ)还原酶上的砷结合位点[40].另外,在研究藻类中砷的生物累积时也发现加入外源物质纳米二氧化钛可通过影响细胞表面的疏水性、细胞膜的完整性和迁移率来影响砷在藻类中的累积[41],加入纳米二氧化钛可显著促进淡水藻类铜绿微囊藻(Microcystis aeruginosa)中As(Ⅴ)的累积和斜生栅藻(Scenedesmus obliquus)中As(Ⅲ)的累积[42]. 作为水生生态系统的重要组成部分,藻类在海洋生态系统砷循环中扮演重要角色,能累积高浓度的无机砷,可以极大降低环境中的砷浓度,因此纳米离子与砷污染物共同累积砷的研究需要更多的关注,以发现更多水生植物对砷的高效解毒途径.水产品由于味道鲜美,营养丰富,同时作为人类优质蛋白的重要来源,深受消费者的青睐,而水产品由于自身易累积重金属的特点,经食物链传递累积至人体中,易造成潜在健康危害,因此有大量对水产品中重金属污染情况并做出膳食健康评价的研究,表1为国内外部分地区近10年来对不同水产品中砷的累积监测情况.表 1 国内外部分地区水产品含砷情况Table 1 The concentration of Arsenic in aquatic products around the world采样年份Year地区Region检测项目Detection projects水产品种类Aquatic products category含量/(mg·kg−1)Concentration参考文献Reference2010—2013茂名市(中国)无机砷Inorganic Arsenic鱼类Fish ND—0.071[43]甲壳类Crustaceans ND—0.073双壳类Bivalves ND—0.040无壳类 Shellless ND—0.1202012巴伦支海Barents sea总砷Total Arsenic红帝王蟹Paralithodes camtschaticus10.000±5.000[44]2015韩国Korea总砷Total Arsenic头足类动物Cephalopods 2.620—13.100[45]1072环 境 化 学43 卷续表 1采样年份Year地区Region检测项目Detection projects水产品种类Aquatic products category含量/(mg·kg−1)Concentration参考文献Reference2015绍兴市(中国)总砷Total Arsenic淡水甲壳类Freshwater crustaceansND—0.710[46]淡水鱼类Freshwater fishND—0.740海水甲壳类Marine crustaceans0.042—7.300海水双壳类Marine bivalvesND—1.510海水头足类Marine cephalopodsND—4.260海水鱼类Marine fish0.028—3.9002016—2017浙江省(中国)无机砷Inorganic Arsenic贝类Bivalves<0.100[47]虾类Shrimp<0.100鱼类Fish<0.100头足类Cephalopods<0.1002017保定市(中国)总砷Total Arsenic淡水鱼Freshwater fish0.024—0.250[48]河虾River shrimp0.180—0.525河蟹River crab0.660—2.6802017德比湖(波兰)Lake Dabie (Poland)总砷Total Arsenic中华绒螯蟹Eriocheir sinensis0.031—0.191[49]2018上海市(中国)总砷Total Arsenic淡水鱼类Freshwater fish0.005—0.474[50]海水鱼类Marine fish0.035—13.000淡水甲壳类Freshwater crustaceans0.052—0.905海水甲壳类Marine crustaceans0.010—6.580头足类Cephalopods0.021—0.475双壳类Bivalves0.020—0.4722018孟加拉国Bangladesh总砷Total Arsenic滇西低线鱲Barilius barila0.083±0.023[51]剑鲑口波鱼Salmostoma acinaces0.021±0.001印度小鳞鲥Gudusia chapra0.278±0.074露斯塔野鲮Labeo rohita<0.020印度细齿鲱Corica soborna0.161±0.037剑鳠Sperata aor<0.0202018图苏库缇(南印度)Thoothukudi, (South India)总砷Total Arsenic甲壳类 Crustaceans 3.190—16.500[52]头足类Cephalopods 1.100—9.1902017—2020广西省(中国)无机砷Inorganic Arsenic鱼类 Fish ND—0.276[53]虾 Shrimp ND—1.141螺 Screw ND—0.150蟹 Crab ND—1.513贝类Bivalves ND—0.138 ND:Not Detected.水产品是我国重点出口的产品,由于所处的自然环境、养殖使用的化学物质及自身特性易累积重金属,经人类摄入造成食品安全问题,因此世界各国组织十分重视水产品中重金属污染物含量问题,出台了相关技术控制法规和标准,如国际食品法典委员会(Codex Alimentarius Commission,CAC)、欧盟、美国、日本、韩国、澳大利亚等都对水产品中重金属污染物出台相关标准和技术法规[54]. 全球各地水生生物生长的水域环境不同,各国消费水产品的情况不同,砷限量值也存在差异,CAC、欧盟、日本、韩4 期冉茂霞等:砷在水生生物中的生物累积、转化及在其他生物体内的代谢毒理学研究进展1073国、孟加拉国等均未规定水产品中总砷/无机砷限量值,而澳大利亚规定鱼类、甲壳类、软体动物无机砷限量值分别为2.0、2.0、1.0 mg·kg −1,国内对砷限量值参照GB 2762—2017《食品安全国家标准 食品污染物限量》,规定水产动物及其制品(鱼类及其制品除外)、鱼类及其制品的无机砷含量分别不能高于0.5 mg·kg −1、0.1 mg·kg −1 [55],印度尼西亚则对海藻及鱼类及其制品的总砷做出最大限量规定,分别为1.0 mg·kg −1和2.0 mg·kg −1[56],美国对甲壳类和贝类中总砷限量值分别不得超过76 mg·kg −1和86 mg·kg −1(表2). 相比澳大利亚及美国的标准,中国对水产品中砷限量更严. 大量的基础科学研究表明,砷在不同水生生物中的赋存形态不同,产生的毒性作用也不同,而国外大部分国家却未对砷做出限量规定,对于各国砷限量差异问题,笔者认为有必要继续完善水产品中砷限量指标,不仅要根据各类水产品生长环境及其自身特性进行研究评估,还要继续研究不同水生生物的砷毒理学效应,有必要细化水产品分类,根据各类水产品中无机砷与总砷占比情况,毒性特点设定更加科学合理的砷限量值.表 2 部分组织或国家砷限量比较Table 2 The comparison of arsenic limited value in some organizations or countries 组织或国家Organizations or countries水产品类别Aquatic products category 限量值/(mg·kg −1)Limited value 参考文献Reference 总砷*Total Arsenic 无机砷**Inorganic Arsenic 孟加拉国Bangladesh———[51]CACCodex Alimentarius Commission———[54]欧盟European Union———日本Japan———韩国Korea———澳大利亚Australia 鱼类Fish — 2.000甲壳类Crustaceans— 2.000软体动物Mollusk —1.000中国China 水产动物及其制品Aquatic animals and their products —0.500[55]鱼类及其制品Fish and fish products —0.100印度尼西亚Indonesia 海藻Seaweed 1.000—[56]鱼类制品Fish products 2.000—美国the United States of America 甲壳类Crustaceans 76.000—[57]贝类Bivalves 86.000— 注:“—”表示该国家或组织未对水产品中总砷及无机砷进行限量规定;*以总砷计,**先测定总砷,若总砷含量水平超过无机砷限量值则需测定无机砷. Note:"-" indicates that the countries or organizations do not set the limited value of total Arsenic and inorganic Arsenic in aquaticproducts; *measured as total Arsenic, ** measured total Arsenic first, if the total Arsenic content level exceeds the limited value of inorganic Arsenic, then measured inorganic Arsenic.1.2 砷在水生生物中的生物转化水生生物易累积砷,并造成一定的毒性作用,环境中的砷进入水生生物体内,经转化代谢作用降低了砷的毒性. 在研究水生生物中砷的生物转化时发现,水生生物对砷的生物转化包括无机砷的生物转化和有机砷的生物转化,水生动物主要吸收无机砷,随后将无机砷氧化或还原,或者通过甲基化生成有机砷形式[58]. 在砷的生物转化过程中,无机砷化合物被细菌、藻类、真菌和人类甲基化,并在甲基化过程中产生甲基化产物MMA 和DMA ,这些甲基化砷是慢性砷暴露的最终代谢物和标志物,转化过程如下:As (Ⅴ)→As (Ⅲ)→MMA Ⅴ→MMA Ⅲ→DMA Ⅴ[59]生物甲基化是无机砷的一种解毒过程,其终产物为甲基化的无机砷,如MMA Ⅴ和DMA Ⅴ,可通过尿液排泄,MMA Ⅲ不能经由尿液排出体外,而是作为一种中间产物留在体内,其具有比其他无机砷更高的毒性,这可能是砷致癌的原因[60].1074环 境 化 学43 卷4 期冉茂霞等:砷在水生生物中的生物累积、转化及在其他生物体内的代谢毒理学研究进展1075一些研究发现,经As(Ⅴ)暴露后的鱼体组织中能检测到更多经As(Ⅲ)直接暴露后的As(Ⅲ),因此,推测As(Ⅴ)能大量转化为As(Ⅲ),并广泛分布在鱼肌肉中. 而As(Ⅴ)的强还原能力可解释在实验暴露结束后鱼类组织中并未检测到As(Ⅴ)的原因,这表明,大量的As(Ⅴ)可能在鱼肌肉中分布前就已经转化为As(Ⅲ). 除此之外,As(Ⅲ)转化为AsB的能力高于As(Ⅴ)[21],因此,As(Ⅴ)还原为As(Ⅲ)可能是水生生物的一种解毒过程. 尽管As(Ⅲ)的毒性强于As(Ⅴ),但是As(Ⅴ)在转化为As(Ⅲ)后,As(Ⅲ)与肽结合、转化为低毒甚至无毒的有机砷, 以及经细胞排出过程中都能降低As(Ⅲ)的毒性. 另外,少数砷形态可能通过其他方式解毒,如通过与金属硫蛋白的结合以及一些甲壳类动物的退壳行为、遗传行为[61 − 62].砷在水生生物体内的生物转化一直是关注的热点,水生生物可以将As(Ⅴ)还原为As(Ⅲ),随后甲基化为MMA和DMA,机体体内存在多种对砷转化代谢过程,如细胞的排泄作用、退壳行为、遗传行为、与机体内的各种金属肽结合、转化为无毒或毒性更低的有机砷. 图2为砷在生物体内的累积转化代谢过程.图 2 砷在生物体内的累积转化代谢过程Fig.2 The process of Bioaccumulation and Biotransformation of Arsenic in organisms2砷在不同生物体中的代谢毒理学作用(Metabolic toxicological effects of arsenic in different organisms)2.1 砷对水生生物体的毒理学作用砷易累积在水生生物体内,参与全球水生生态系统中的砷循环,由天然形成和人类生产过程中产生的砷易累积在水生生物体内,可通过改变机体内的各种酶和代谢物,导致机体发生组织、代谢上的紊乱[63],既能降低砷对机体的毒性又能导致机体遭受更大的毒性,并且不同的砷形态对不同水生生物产生的毒性不同. 例如,Rahman等[64]对小球藻(Chlorella sp. strain CE-35)、浮萍(Lemna disperma)、水蚤(Ceriodaphnia cf. dubia)的3种淡水生物进行As(Ⅲ)、As(Ⅴ)和DMA毒性大小比较,结果发现As(Ⅲ)对浮萍的毒性最大,对小球藻的毒性最小,As(Ⅲ)、As(Ⅴ)对水蚤的毒性相当,DMA对浮萍和水蚤的毒性最小. 另外,环境中的磷浓度、pH值、盐度和溶解有机物(DOM)水平等均会影响砷对水生生物的毒性[62]. He等[65]研究不同pH下明亮发光杆菌(Photobacterium phosphoreum)、大型水蚤(Daphnia magna)、斑马鱼(Danio rerio)的3种水生生物中的砷毒性,发现不同pH下这3种水生生物受到砷毒性的大小随pH的变化存在生物特异性. Rahman等[64]对3种淡水浮游生物的毒性试验中发现,As(Ⅲ)和As(Ⅴ)对小球藻和浮萍的毒性与培养基中磷酸盐(PO43-)浓度有关,小球藻具有将As(Ⅴ)还原为As(Ⅲ)的能力,这种途径可能是小球藻对砷的耐受机制,以降低As(Ⅴ)对机体的毒性.由于生物体通过磷酸盐转运体吸收As(Ⅴ),通过甘油蛋白吸收As(Ⅲ),因此不同的转化途径可能导致不同的砷毒性[66]. 尽管砷对淡水浮游植物的毒性因浮游植物种类的不同而有极大差异,但大部分毒性生物测定表明,As(Ⅴ)比As(Ⅲ)对淡水浮游植物的毒性更大,而As(Ⅲ)对海洋浮游植物毒性更大. 尽1076环 境 化 学43 卷管As(Ⅲ)和As(Ⅴ)在淡水系统中占主导地位,但砷形态对水生生物的毒性由于与水中磷酸盐等化学物质相互作用而变得更为复杂,这些化学物质会影响As(Ⅴ)的生物利用度和吸收[39],而As(Ⅲ)具有更高的生物利用度,因此,As(Ⅲ)比As(Ⅴ)对海洋浮游植物的毒性更大.在淡水和海水中,水生生物体内的主要砷形态为AsB,这表明水生生物为保护自身机体免遭更大的毒性作用,可以高毒的无机砷最终转化为几乎无毒的AsB. 另外,不同形态的砷对不同的水生生物具有不同的毒性,对于砷产生毒性差异的原因可能是由于生物体自身因素,如摄取砷的途径、代谢途径及机体自身对重金属的解毒机制上的差异性,以及非生物因素,如砷的暴露浓度、暴露时间、暴露介质中的pH、PO43-水平等.2.2 砷在小鼠/大鼠体内的代谢毒理学作用2.2.1 砷在小鼠/大鼠体内的代谢砷在全球水生生态系统内循环,通过食物链传递,产生生物放大作用,对不同的生物体产生不同的毒性,鱼类、甲壳类、双壳类水生动物易累积砷,通过食物链传递,最终累积到人体中,对人类健康造成了潜在危害,因此,砷对人体的代谢毒性受到了广泛的研究. 为了解膳食摄入砷对不同生物体的毒理学作用,研究者们通常采用小鼠/大鼠模拟不同生物体体内砷代谢情况,测定小鼠/大鼠各器官中砷形态,同时收集它们的排泄物以反映砷的代谢毒理学效应. 例如,王旭等[67]对81只大鼠进行4种砷形态(As(Ⅲ)、As(Ⅴ)、MMA、DMA)为期28 d的动物代谢实验,发现这4种砷形态的主要排出方式为粪便和尿液,少部分仍保留在血液中. 赵梦醒等[68]研究海带中砷元素在大鼠消化系统和血液中的形态变化,发现随着海带中砷形态在大鼠体内代谢,除大鼠血液中的砷形态无变化外,海带中的砷形态在消化系统中的形态和含量均发生变化. 于霄云等[69]探讨外源性谷胱甘肽(GSH)和亚硒酸钠对饮水砷暴露小鼠肝、肾和血中砷代谢的影响,发现GSH组小鼠肝中和血液中DMA含量和总砷含量均高于对照组,GSH组小鼠肝中砷一甲基化率和二甲基化率与砷组小鼠对应的甲基化率比较升高,因此,对小鼠给予外源性GSH可以促进无机砷在小鼠体内甲基化代谢.慢性砷暴露会导致小鼠代谢紊乱,如葡萄糖代谢受损和能量消耗减少,在慢性砷暴露对小鼠皮层和血清代谢组学的影响实验中发现,慢性砷暴露可通过干扰皮层和血清氨基酸类代谢和三羧酸循环、神经递质合成障碍以及干扰能量代谢而影响中枢神经系统功能[70 − 71]. 另外,亚慢性砷暴露对小鼠大脑组织神经递质代谢酶及其受体基因表达谱有所影响,对小鼠进行As2O3和锑(Sb)的共同暴露会导致小鼠氧化应激、肝组织脂质代谢紊乱和脂肪变性,这可能是肝组织中线粒体吞噬和线粒体通路凋亡的重要驱动因素,而这种线粒体吞噬和凋亡极大程度上诱导细胞死亡[72]. As2O3可能导致小鼠大脑组织部分神经递质的合成和分解代谢酶基因表达下调,且干扰其部分受体基因的表达[73]. 在对小鼠进行慢性饮水无机砷暴露时发现小鼠体内无机砷代谢产物的蓄积与分布具有明显的组织和区域特异性,由于无机砷代谢产物具有不同的毒性,推测膀胱和脑组织可能是慢性无机砷暴露发挥毒性最重要的靶器官,而详细机制仍需进一步探讨[74]. 目前众多研究选择小鼠/大鼠作为砷形态致毒机制研究的主要对象,而其准确致毒机制仍然不是十分清楚,虽然小鼠/大鼠常被用做毒理学研究,但由于砷在生物体内的代谢物及砷本身都会与细胞内外大分子发生作用,且小鼠/大鼠与人体内的的砷代谢机制仍有很大差别[75],因此,未来对砷代谢机制仍然需要大量研究.2.2.2 砷在小鼠/大鼠体内的毒性小鼠/大鼠组织器官中的砷经代谢后极大降低了砷对机体的毒性,目前研究者对砷在小鼠/大鼠体内的毒性研究取得了一定的成果,无机砷已被列入Ⅰ类致癌物,对人体具有一定的致畸致癌性,而有机砷则被认为毒性较小,但大量的研究发现小鼠/大鼠中的砷经代谢后,器官组织中能检测到大量的DMA,而DMA作为无机砷的主要代谢产物本身具有独特的毒性,研究发现在小鼠和大鼠的肺以及人肺细胞中的DMA会诱导器官特异性损伤,造成DNA单链断链,造成这种损伤的主要原因是DMA的过氧自由基和肺内组织产生活性氧[76 − 77],DMA染毒后可使小鼠骨髓细胞有丝分裂指数明显上升,造成DNA单链断裂,而DMA的进一步代谢产物二甲基砷化物能与氧分子反应产生活性氧,使得DNA链上的C8位鸟嘌呤碱基被羟基自由基和单线态氧攻击生成8-羟基脱氧鸟苷(8-OHdG),导致DNA氧化损伤[78]. 虽然DMA是有机砷形态,其毒性远低于无机砷毒性,但仍然不能忽视其毒性作用.。
砷中毒生物标记物的研究进展

可在病情发展至中、晚期显示异常,SBA、Y—GT、GST、ET联合应用在砷致肝损伤的筛检及 动态观察其病情发展中有重要参考价值。对砷中毒患者肾功能检测发现,在常规肾功能检查 正常的情况下,可见血、尿B z微球蛋白(U B z—MG)、尿白蛋白(UALB)、尿N-乙酰一B—D一 氨基葡萄糖苷酶(NAG)异常,提示砷可在早期致肾脏损害。而作为“肿瘤标志物”的血清 铁蛋白(FP)、唾液酸(SA)、岩藻糖苷酶(AFU)的改变在砷致癌进程中有一定预警意义。 虽然这些指标缺乏特异性。但在明确砷接触且排除其他病因的前提下,仍不失为有效且易于 检测的效应标志物。 2.2 8一羟基一2脱氧鸟苷(8一伽dG)DNA修复的产物无需进一步代谢即排人尿中,因此许多研 究都试图通过分析尿中DNA加合物或DNA氧化损伤修复产物等生物学标志物,对体内氧化负 荷进其是由活性 氧(ROS)攻击核酸的产物,可进~步导致突变、癌变等发生。
hGST0卜1是MMA的还原酶,可以使M(V)转化成MMA(Ⅲ),属于谷胱甘肽转移酶的亚目。
Tanaka等心51认为hGST0卜1两种变异体A1a140Asp和Thr217Asn在生物活性方面的不同可能是 个体对氧化应激和对无机砷易感性差异的原因。虽然目前缺乏可信的资料阐明GST基因与地 方性砷中毒的关系,但可以肯定,其基因多态性在不同种群中的分布频率差异可能是构成人 群间易感性差异的遗传基础。 3.2髓过氧化物酶(船o) MPO是主要存在于嗜中性粒细胞和单核细胞中的一相代谢酶。能 使骨髓细胞生成熟分化过程中产生氧自由基,引起DNA损伤,并能产生次氯酸,参与前致癌
采用横断面调查研究儿童砷暴露与DNA氧化损伤的关系,结果表明,尿砷含量高的儿童 尿8—0HdG水平明显高于尿砷含量低的儿童…。由于8—0HdG影响因素少,检测方法敏感。因 此,可广泛应用于砷中毒的早期监测。 2.3遗传毒效应指标外源化合物遗传效应最常用的3个检测终点是微核率(心)、姐妹染色 单体互换(SCEs)、染色体畸变(cAs)。唧是目前最常用的反映细胞遗传物质损伤的指标之一。 对西班牙高砷饮水(0.75mg/L)地区106名居民和111名健康对照的研究表明,高砷暴露诱发外 周血淋巴细胞州率明显高于对照组旧1。Mahata等旧。比较了饮用高砷井水居民细胞遗传学改 变,饮水中砷浓度(211.70±15.28)垤/L地区,发砷、指(趾)甲砷和尿砷分别 (9.04±0.78)№/g、(5.63±O.38)№/g和(140.52±8.82)垤/L的59名居民外周血淋巴细胞 scEs互换率为7.26/细胞,明显高于饮水中砷浓度为(6.35±0.45)№/L、年龄和社会经济状 况匹配的发砷、指(趾)甲砷和尿砷分别为(0.44±O.03)腿/g、(0.30±O.02)№/g和 (5.91±O.49)№/L的36名健康居民SCEs互换率(5.95/细胞)。体外研究表明u训,砷低浓度时, 染色体畸变类型主要以染色单体断裂为主;砷浓度为200|l g/L时,染色单体及染色体断裂明 显增加的同时,染色单体互换开始出现;砷浓度达400 u g几时,染色单体互换率与对照组的 差异有显著性(尸<0.05)。染砷浓度与细胞畸变率、染色体畸变出现的频度以及染色体畸 变类型的出现有明显的剂量一效应关系。 2.4脂质过氧化(LP) 脂质过氧化(LP)是指机体通过酶系统和非酶系统反应产生的氧自由 基,攻击生物膜磷脂中多不饱和脂肪酸和脂肪酸而引起的一系列氧化过程。近年来,脂质过 氧化学说在阐述砷的毒作用机制上引起了关注,动物亚慢性毒性实验结果表明u¨,随着砷染 毒剂量的增加,小鼠血清及肝脏组织中丙二醛(肋A)和谷胱甘肽S一转移酶(GSTs)含量显著增 加;超氧化物歧化酶(SOD)、谷胱甘肽过氧化物酶(GSH—Px)活性显著降低,且呈剂量~效应 关系。 2.5金属硫蛋白(MT) MT是富含半胱氨酸的低分子蛋白质,具有调节内源性金属的平衡、对 重金属污染解毒、清除自由基、参与应急反应作用。因此,MT是某些重金属接触生物标志物 和早期生物学效应的标志物。已有研究发现砷中毒小鼠肾脏MT表达增高u副。MT可以显著减弱 砷对DMA的基因毒性,因此,MT是一种有效的砷毒性拮抗剂、抗氧化剂和砷的化学致癌保护 剂n 3‘。但在贵州燃煤型病区检查砷中毒病人口腔黏膜脱落细胞中检测到MT—l基因表达显著较 低u制。上述研究提示,人体MT表达可能存在差异,其原因有待进一步研究。 2.6应激蛋白或热休克蛋白(HsP) 应激诱导细胞产生一组高度保守的蛋白质,被命名为热 休克蛋白(HSP)。HSP参与多种蛋白的折叠、装配及转运等蛋白质加工修饰过程。HSP在细胞 周期调节、DNA损伤修复、基因产物代谢等过程中起重要作用,因此与细胞转化和恶变过程 密切相关。在物理和化学因素刺激作用下,机体可被诱导高度表达HsP。砷暴露引起多种热 休克蛋白高度表达,体外研究结果表明,亚砷酸钠无论是急性作用还是较长时间作用于人膀
水产品中砷的质量控制研究进展

水产品中砷的质量控制研究进展水产品中砷的质量控制研究进展? 戴文津1,3,杨小满2,3,陈华1,3,孙恢礼1,3 摘要:水产生物资源的开发利用一直是生物资源可持续发展研究的重要部分,但某些水产食品于养殖捕捞水域环境质量劣化、自身易富集重金属的原因,含砷量较高,不仅具有潜在的食用危险性,也限制了其作为加工原料的使用,因此对水产食品中无机砷进行检测并减低其砷含量尤为重要。
就水产品中无机砷的分析检测技术、含量控制方法以及砷脱除技术研究现状和发展前景进行了总结和展望,并提出具有较好应用前景的水产品脱砷方法。
关键词:水产品;砷;检测;质量控制;脱除中图分类号:S912文献标识码:A 文章编号:1004-874X11- Advances in analysis and content controlof arsenic in seafood Dai Wen-jin 1,3, Yang xiao-man2,3, Chen Hua 1,3, Sun Hui-li 1,3 Abstract:Development and utilization of living aquatic resources is an important part of the sustainable development of living resources. However some aquatic foods are rich in arsenic because of cultivation and fishing environment deterioration and the biological enrichment of heavy metal. They are potentially hazardous, and their uses as raw material are also limited. So analyzing and reducing the arsenic contents in seafood become very important. This paper represented the research status and development trends of analysis, content control and removal method of arsenic in seafood. And some promising ways were also put forward in the paper. Key words:aquatic food;arsenic;analysis;quality control;removal 水产品中砷存在形态复杂多样,主要有二甲基胂酸盐、甲基胂酸盐、无机砷[包括As(Ⅲ)和As(Ⅴ)]等,其中致毒性和致癌作用主要取决于无机砷。
海产品中的砷及其代谢机制的研究进展

品中砷的含量与其所在水环境以及饮食习惯有很大关 并可使心脏及脑组织缺血,引起虚脱,意识消失及痉
系。生活在海洋底部的鱼(如比目鱼)所含砷的浓度 挛等。砷被吸收后,可麻痹血管运动中枢,直接作用
较高;生活在海洋地表的贝类也有较高浓度的砷[27]; 于毛细血管,使脏器的微血管发生麻痹、扩张和充血,
长期生活在砷浓度较高的海水中的鱼摄入了高浓度的 以致血压下降;并且被人体吸收的砷,部分贮留于肝
DM A
0.6
weight)
AsB
5.4
总类 国家
砷形态
含量
参考文献
AsC
<LOD
褐藻
挪威
中国
总砷
1220
[16]
DM AA
0.013
DM AP
<LOD
DM AE
0.005
总砷
13.1-33.8
(As(V))、砷胆碱(AsC)、砷甜菜碱(AsB)、砷糖、砷脂等。各种形态的砷通过不同代谢途径产生不同的代谢产物,从而表现出不
同的毒性。砷的形态主要是通过高效液相色谱与电感耦合等离子体质谱仪的联用(HPLC-ICP-MS)来测定的。有机砷中砷胆碱(AsC)
的代谢产物为砷甜菜碱(AsB)和砷脂;砷甜菜碱(AsB)经尿液直接排出;不同的砷脂代谢产物不一样;砷糖的代谢产物至少 12
其他海产品中,超过 90%的砷是以毒性较小的有机形 织和体液中。所以,水体、食物和大气的砷污染,都
态存在的,无机砷的含量仅占 0.02%~11%[23~25]。海产
会直接危及人畜的健康。在体内,砷可与细胞内巯基
品中砷的总量,受海产品种类及所在地区影响。中国 酶结合而使其失去活性,从而影响组织的新陈代谢,
集,特别是以水体中浮游生物为食物链的水产生物体 量。
原子荧光法测定海产品中总砷前处理方法的研究

的紫菜 , 贝 质 控 品 GB 1 0 4 地 球 物 理 地 球 化 学 扇 w O2( 勘 查研究 所提 供 ) 。 12 仪 器 . AF 一 2 0 S 3 E原 子 荧 光 分光 光 度 计 , 特 梅
勒 AG2 5电 子 天 平 , 4 MARS 5微 波 消 解 仪 ,MI L — LI
l 及 1 0g氧化 镁 , 匀后 置 于 电热 板上 炭化 至无 0ml . 混 黄色 烟 , 人 5 0C马 沸 炉 中灰 化 4h 移 5。 。取 出冷 却 , 小
成 特点不 同 , 品 的前处 理 目前 有 湿法 消 解 、 灰 化法 样 干
和微波消解 法等数 种『 , 2 现试通 过 比较 不 同的样 品前处 ] 理 方法 , 来评价样 品前处理对 海产 品总砷 测定 的影 响 。
氢氧 化钠用 水溶 解定 容 至 50ml2 硼 氢化 钾 : 取 0 ; 称
1 氢化 钾溶 于 5 0ml / 0g硼 0 g L氢 氧 化钠 溶 液 中 ; 2 / 盐
酸溶液 ( o . 5 吸取 5 超 纯 盐 酸用 水 定 容 至 10 0 0) 0ml 0 ml10 g ml ;0 0M / 砷标 准溶 液 ( 国家 标 准物 质 研 究 中心
1 5 3 微 波 消解 法 ( 波 法 ) 称 取 0 1 0 3g样 品 .. 微 : . - .
11 样 品 .
虾 粉 ( 力 验 证 T 4 2样 品 ) 市 场 购 买 能 04 ,
于高 压消 解罐 内, 加入 6ml 酸 , 过氧 化氢 , 硝 2ml 放置 过 夜后 于 消解仪 内消解 ( n升至 10C保 持 5 n 5mi 2 ̄ , mi 5mi 至 10C保 持 5 mi , n升 至 1 5 n升 5。 n 5mi 8 。 持 C保 1 i) 5r n 。消解 完后 放加 热板 上赶 酸 . a 备用 。
HPLC-AFS联用测定海产品中砷的形态

5 3 0 0 1 1 ,C h i n a ;3 . S y ma n t e c S o t f wa r e D e v e l o p me n t ( C h e n g d u ) C o . L t d, C h e n g d u 6 1 0 0 4 1 , C h i n a ;4 . G u a n g x i Bo t a n i c a l Ga r d e n o f
[ 1 . Gu a n g x i C e n t e r o f An a l y s i s a n dT e s t Re s e a r c h , Na n n i n g 5 3 0 0 2 2 , C h i n a ; 2 . Na n n i n gC o n s t r u c t i o n a n dI n s t a l l a t i o nC o . L t d , Na n n i n g
HP L C - AF S联用测定海产 品中砷 的形态
刘守 廷 , 蒋 天成 。 李健 梅 , 罗平 , 陶弋 , 宋业 成
[ 1 . 广西分析测试研究 中心 , 南宁 5 3 0 0 2 2; 2 . 南宁市建工建筑安装有 限公司 , 南宁 5 3 0 0 1 1 ; 3 . 赛门铁克软件开发( 成都 )
有限公 司, 成都 6 1 0 0 4 1; 4 . 广西药用植 物园, 南宁 5 3 0 0 2 3 ]
摘要
建立 了高效液相 色谱 一原子 荧光分光光度 法测定海产品 中无机砷 ( As V, As I I I ) 、 有机砷( DMA, MMA,
As B) 含 量 的方 法。样 品 经含 l 0 %( 体积 分数 ) HC 1 的提 取 液振 荡提 取 、 离心 分 离、 二路 形 态分 析预 处理、 高效液
砷在海洋食物链中的生物放大潜力及发生机制探讨

生态毒理学报Asian Journal of Ecotoxicology第14卷第1期2019年2月V ol.14,No.1Feb.2019㊀㊀基金项目:国家自然科学基金面上项目(41876133),国家重点基础研究发展计划项目(2015CB452904)㊀㊀作者简介:杜森(1992-),男,博士研究生,研究方向为海洋生态毒理学,E -mail:dusen14@ ㊀㊀*通讯作者(Corresponding author ),E -mail:zhangli@DOI :10.7524/AJE.1673-5897.20181112002杜森,张黎.砷在海洋食物链中的生物放大潜力及发生机制探讨[J].生态毒理学报,2019,14(1):54-66Du S,Zhang L.Biomagnification potential and the mechanisms of arsenic in marine food chains [J].Asian Journal of Ecotoxicology,2019,14(1):54-66(in Chinese)砷在海洋食物链中的生物放大潜力及发生机制探讨杜森1,2,张黎1,*1.中国科学院南海海洋研究所,中国科学院热带海洋生物资源与生态重点实验室,广州5103012.中国科学院大学,北京100049收稿日期:2018-11-12㊀㊀录用日期:2019-01-09摘要:砷是世界范围内危害最大的环境污染物之一,也是近海区域一种常见污染物㊂本文综述了近年来砷在海洋生态系统中累积㊁转化及传递的最新研究进展㊂海洋生物普遍具有较高含量的砷,这些砷主要为低毒性的有机砷形态㊂砷在许多海洋食物链/网中被生物放大,造成高营养级生物中的砷富集,可对生物与人类健康产生潜在危害;这与砷在淡水食物链/网中普遍被生物减小的现象形成鲜明对比㊂海洋鱼类和贝类等生物可将吸收的无机砷通过生物转化合成砷甜菜碱等有机砷形态,而有机砷比无机砷具有更高的食物链传递能力,可导致海洋鱼类富集更高浓度的砷㊂因此,砷在海洋生物中的有机形态可能有助于砷沿着海洋食物链/网富集,在某些情况下被生物放大㊂今后应该加强对不同砷形态在海洋食物链/网中传递及相应影响因素的研究,并通过室内模拟实验与野外调查相结合进行验证,从而加深对砷的生态毒理和生物地球化学作用的科学认识,对准确评估预测砷的生态风险和保障海洋生态安全有重要意义㊂关键词:砷;食物链;海洋;生物放大;形态文章编号:1673-5897(2019)1-054-13㊀㊀中图分类号:X171.5㊀㊀文献标识码:ABiomagnification Potential and the Mechanisms of Arsenic in Marine Food ChainsDu Sen 1,2,Zhang Li 1,*1.Key Laboratory of Tropical Marine Bio -resources and Ecology,South China Sea Institute of Oceanology,Chinese Academy of Sci -ences,Guangzhou 510301,China2.University of Chinese Academy of Sciences,Beijing 100049,ChinaReceived 12November 2018㊀㊀accepted 9January 2019Abstract :Arsenic (As)is one of the most hazardous environmental pollutants and widely exists in the coastal area.This paper reviews the bioaccumulation,biotransformation and trophic transfer of As in marine ecosystems.Marine organisms usually accumulate high level of As mainly in low -toxic organic species.As could be biomagnified a -long some marine food chains/webs,resulting in the high bioaccumulation in higher trophic levels and potential harm to these organisms.In contrast,As is commonly biodiminished in freshwater food chains/webs.Marine fish and shellfish could biotransform inorganic As to organic species,especially to arsenobetaine,which have higher trophic transfer ability and then cause high As bioaccumulation.As biomagnification is probably related to the high第1期杜森等:砷在海洋食物链中的生物放大潜力及发生机制探讨55㊀content of organic As species in marine organisms.The future research should pay more attention to the biomagni -fication potential of specific As species by the combination of field investigation and indoor experiments.It will be helpful for the better understanding of the ecotoxicology and biogeochemistry of As,the assessment of the ecologi -cal risks of As,and the safeguard of the marine ecological safety.Keywords :arsenic;food chain;marine;biomagnification;speciation ㊀㊀砷(As)是一种广泛分布于自然界中的有毒类金属元素,是危害最严重的环境污染物之一㊂砷位于元素周期表中第四周期第五主族元素,主要以+3和+5的亚砷酸盐和砷酸盐的形式存在于环境中㊂砷主要用于杀虫剂㊁除草剂㊁杀真菌剂和土壤消毒剂,也用来除莠病和消毒种子;工业中也常用来有色金属的冶炼㊁半导体元件制造业和净化工业气体等[1-2]㊂在综合考虑毒性大小和污染发生频率的基础上,美国有毒物质与疾病登记署(Agency for Toxic Substances and Disease Registry)和环境保护署(Envi -ronmental Protection Agency)将砷排在2017年 优先污染物列表 的首位[3]㊂不同来源的砷通过陆地径流㊁大气沉降和废水直接排放等途径进入海洋生态系统,导致砷污染成为近海区域严重的环境问题之一[4]㊂海洋生物普遍具有较高的砷累积和代谢转化能力,因而在体内含有高浓度的砷,并主要以有机砷形态存在㊂这导致海产品成为许多国家和地区人群砷摄入的主要来源(在美国和日本超过90%),一直受到密切关注[5]㊂食物相暴露是海洋捕食者(像腹足类和鱼类等)累积痕量元素的主要途径[6],这导致痕量元素沿食物链进行传递㊂污染物是否具有生物放大(biomag -nification)的能力,是评估其对环境造成危害的重要依据之一㊂具有生物放大能力的污染物在沿食物链/网传递的过程中,会随营养级的增加在生物体内逐级富集,在高营养级生物中达到较高水平,对生物与人类健康产生潜在危害,因此其生态风险和人类健康风险就应当受到格外的关注㊂有机汞以及持久性有机污染物(如多氯联苯)等就是典型的具有高生物放大能力的物质,几十年来已被众多研究所关注[7-8]㊂砷通常不被认为是具有生物放大能力的污染物,过去的研究发现,砷在淡水和陆地食物链/网中确实常被生物减小(biodiminution)[9-10]㊂然而,近些年不少研究发现,砷在某些海洋食物链/网中出现了生物放大的现象㊂迄今为止,关于砷沿食物链/网传递的研究却非常匮乏,而且对相关机理的认识也不明确㊂因此,本文总结了近20多年来国内外有关砷在典型海洋生态系统中食物链/网传递作用的相关研究,初步探讨砷在海洋食物链/网中是否具有生物放大潜力以及相关砷在不同食物链/网中传递作用的关键核心与影响因素,有助于加深对砷的生物有效性和生物地球化学行为的认识,对准确评估预测砷的生态风险和保障海洋生态安全有十分重要的意义㊂1㊀砷在海洋生态系统中的浓度与形态(Arsenic concentration and speciation in marine ecosystems )砷具有不同的化学形态(如图1),不同形态砷的生物毒性具有很大差异㊂砷在海水和沉积物环境中图1㊀海洋生物体内常见砷化合物的结构Fig.1㊀Structures of common arsenic species in marine organism56㊀生态毒理学报第14卷主要以无机形态存在,而在海洋生物中已鉴定出20多种不同的无机和有机形态,其中无机砷主要包括高毒性的三价砷[As(III)]和五价砷[As(V)],有机砷包括低毒的甲基砷(Monomethylarsonate,MMA)㊁二甲基砷酸(Dimethylarsinate,DMA)㊁三甲基砷(Trimethy-larsine,TMA),微毒的砷糖(Arsenosugars,AsS)㊁砷胆碱(Arsenocholine,AsC)和砷甜菜碱(Arsenobetaine,AsB)等[11-13]㊂与大部分重金属通过络合作用形成各种形态不同,这些有机砷形态为共价键形成的化合物,具有较为稳定的结构,例如砷甜菜碱即三甲基砷乙酸酯(CH3)3As+CH2COO-,由甜菜碱中氮原子被砷原子取代形成[13]㊂目前有关海洋生物中砷的研究主要集中在对不同海洋生物中砷的浓度和形态的野外调查[14-16],有少量研究通过室内砷暴露模拟实验分析海洋生物对环境和食物中砷的累积和转化机制[17-18]㊂野外调查显示,通常海水中砷浓度较低(μg㊃L-1级),但海洋生物中普遍具有高砷水平,在未受污染的区域处于1 ~100μg㊃g-1范围内,在一些污染区域可超过1000μg㊃g-1[19]㊂表1总结了世界范围内海洋生物整体或肌肉组织中总砷浓度范围和已检出的砷化合物形态㊂由于海洋生物组织中总砷浓度差异很大,这里用几何平均值表征其典型浓度㊂在不同类群海洋生物中,海藻和海草中总砷浓度较低[21]㊂海藻中的砷约1%~50%为无机形态,更多的是以砷糖和甲基砷为主的有机形态[20,27],这与淡水和陆地植物中以无机砷为主形成鲜明对比㊂浮游植物与浮游动物处于低等水生食物链,后者常以前者为食,所以在浮游动物中也可以检测到浮游植物中出现的砷形态㊂浮游动物体内的砷形态也间接反映了其摄食食性,即杂食性种类浮游动物体内累积AsB,而食草性浮游动物体内累积砷糖,并将其转化为微量AsB[28-29]㊂海洋无脊椎动物累积的砷浓度普遍较高,而多毛类和腹足类等底栖无脊椎生物具有很高的砷含量(例如马氏独毛虫Tharyx marioni中超过2000μg㊃g-1和细角螺Hemifusus ternatanus中约300μg㊃g-1)[21]㊂多毛类动物沙蠋(Arenicola marina)体内的砷主要为三价砷(58%)和五价砷(16%),有机砷不到20%;对其进行水相As(V)暴露后发现该多毛类能将As(V)转化成As(III)和DMA[30]㊂Chen等[31]对菲律宾蛤仔(Ruditapes philippinarum)进行30d无机砷暴露后发现,鳃部累积的砷显著高于消化腺,蛤体内砷的主要形态为AsB和DMA;亚细胞分布显示,砷主要分布在金属硫因蛋白(metallothionein-like pro-teins,MTLPs)和富金属矿体(metal-rich granules, MRG)中㊂MTLPs能够与金属结合从而在细胞内金属的转运和隔离中发挥重要作用[32],并增加生物对砷的耐受能力㊂海洋鱼类中的砷浓度范围也非常大,与无脊椎动物类似,底栖鱼类比浮游鱼类含有更高的砷浓度(例如孔鰕虎鱼Trypauchen vagina中高达134μg㊃g-1[22])㊂砷甜菜碱是绝大多数海洋动物组织中最主要的砷形态,通常占海洋甲壳类和软体动物组织中表1㊀海洋生物体内总砷的浓度和所检出的砷化合物形态[20-26]Table1㊀The total arsenic concentration and arsenic speciation of marine organisms[20-26]生物类群Biological group砷浓度范围/(μg㊃g-1,干重)As concentration range/(μg㊃g-1,dry weight)砷浓度平均值/(μg㊃g-1,干重)As concentration average/(μg㊃g-1,dry weight)砷形态As speciation藻类Algae0.1~35 4.32As(III)㊁As(V)㊁MMA㊁DMA㊁AsS 海草Seagrass0.16~0.590.28As(III)㊁As(V)浮游动物Zooplankton0.2~24.4 2.10As(III)㊁As(V)多毛类Polychaetes 5.0~273929.19As(III)㊁As(V)㊁MMA㊁DMA㊁AsB 甲壳类Crustacean0.1~270.514.86As(III)㊁As(V)㊁MMA㊁DMA㊁AsC㊁AsB 双壳类Bivalve0.6~21410.44As(III)㊁As(V)㊁MMA㊁DMA㊁AsC㊁AsB㊁AsS 腹足类Gastropods8.0~53351.97As(III)㊁As(V)㊁MMA㊁DMA㊁AsB头足类Cephalopoda 4.0~49.516.11As(III)㊁As(V)㊁MMA㊁DMA㊁AsB浮游鱼类Planktonic fish0.5~7.8 3.28As(III)㊁As(V)㊁MMA㊁DMA㊁TMA㊁AsC㊁AsB 底栖鱼类Benthic fish 5.6~449.519.48As(III)㊁As(V)㊁MMA㊁DMA㊁TMA㊁AsC㊁AsB第1期杜森等:砷在海洋食物链中的生物放大潜力及发生机制探讨57㊀总砷的50%~95%,在海洋鱼类组织中占80~99%以上㊂相反,砷在淡水生物中主要以无机砷为主,虽然也有研究报道砷甜菜碱的存在,但它占总砷的比例远低于海洋生物[24]㊂海洋鱼类累积砷的生物动力学模型可以描述为[33-34]:d C /d t =k u ˑC w +AE ˑIR ˑC f -(ke +g )ˑC式中,d C /d t 为累积速率,k u 为水相吸收常数;C w 为水中砷浓度,AE(assimilation efficiency)为同化率;IR 为摄食率,C f 为食物中砷浓度,k e 为排出速率常数,g 为生长速率,C 为生物体内砷浓度㊂砷可以通过水相和食物相进入海洋生物体内,k u ㊁AE 和k e 是影响海洋生物体内砷累积的3个重要参数,表2列举了通过室内模拟实验得出的常见海洋生物中的生物动力学参数㊂海洋鱼类对水相砷的吸收速率远小于小型底栖生物,尤其是在双壳类中,砷的水相吸收速率和同化率较高,而排出速率相对较低,说明其对砷的累积能力较强,这也与调查中发现的小型底栖生物高砷含量一致㊂引起海洋无脊椎动物和鱼类组织中砷浓度变化的原因尚不完全清楚,但可能与环境条件(温度㊁盐度和磷酸盐等)引起的砷的生物可利用性变化以及物种间差异有关㊂Larsen 和Francesconi [40]发现波罗的海和北海海域鱼类体内总砷浓度(0.04~10.9μg ㊃g -1,湿重)与盐度(8ɢ~32ɢ)之间存在显著的正相关关系(P <0.001)㊂底鳉(Fundulus heteroclitus )对水相砷的浓缩因子(concentration factor,CF)与盐度呈正相关的关系[36]㊂Wu 等[41]发现盐度不仅影响As(V)对菲律宾蛤仔体内氧化应激压力和抗氧化酶系统活性,还干扰渗透压调节和能量代谢㊂磷酸盐(PO 3-4)水平会影响海洋浮游植物杜氏藻(Dunaliella tertiolecta )体内砷的循环[42]㊂虽然目前有关海洋生物砷含量和形态的数据很多,但是由于这些研究通常仅针对一种或数种生物开展,而这些生物又受到各自所处不同环境条件的影响而具有很大的砷浓度范围,因此很难根据这些数据分析不同营养级生物对于砷累积的差异性,只有针对特定生态系统中较完整的食物链进行调查研究,才能反映砷沿食物链传递的特征和规律㊂2㊀砷在海洋食物链/网中的生物放大潜力(Arsenic biomagnification potential along food chains /web )生物放大(biomagnification)一般是指化学物质在生物摄食作用下,导致了生物体内化学物质的浓度高于食物中的浓度的现象㊂也有一些学者将生物放大现象定义为浓度随营养级的增加而升高,当生物放大系数(biomagnification factor,BMF)大于1时,这个物质就被生物放大㊂前者将浓度的升高限制在食物相暴露的因素下,而后者的影响因素包括通过水相暴露和食物相暴露,其中生物从水相吸收污染物的过程称为生物浓缩(bioconcentration)[43]㊂由于海洋动物中的砷累积主要由食物相获得[35-39],所以对于海洋食物链而言,这2种定义下的砷生物放大作用差异不大㊂表3总结了砷在不同类型海洋生态系统中沿食物链传递的数据㊂这些研究较为综合地分析了某一特定生态系统中环境(海水和沉积物)中砷的背景浓度和不同类群生物中的砷含量,同时部分研究通过氮稳定同位素(δ15N)确定了这些生物的营养级,可较有效反映砷在自然状况下的食物链传递行为㊂从表中可以发现,不同类型的生态系统中砷的食物链/网的传递存在明显的差异㊂总体上说,砷在水生生表2㊀海洋生物中砷累积的生物动力学参数Table 2㊀Biokinetics parameters of As bioaccumulation in marine organisms类群Group 种名Speciesk u /(L ㊃g -1㊃d -1)AE/%k e /d -1鱼类Fish 细鳞鯻[35](Terapon jarbua )[35]0.0015 5.50.03鱼类Fish 底鳉[36](Fundulus heteroclitus )[36]0.0006--鱼类Fish 诸氏鲻鰕虎鱼[37](Mugilogobius chulae )[37]0.001918.70.24多毛类Polychaete 沙蠋[38](Arenicola marina )[38]0.16487.80.0478多毛类Polychaete 沙蚕[39](Nereis diversicolor )[39]0.0570.2890.0488双壳类Bivalve蛤[18](Scrobicularia plana )[18]3.954400.023注:k u 为水相吸收常数;AE 为同化率;k e 为排出速率常数㊂Note:k u stands for waterborne uptake rate constant;AE stands for assimilation efficiency;k e stands for efflux rate constant.58㊀生态毒理学报第14卷表3㊀砷沿不同水生生态系统食物链/网传递研究汇总T a b l e 3㊀S u m m a r y o f a r s e n i c t r o p h i c t r a n s f e r i n d i f f e r e n t a q u a t i c e c o s y s t e m s研究区域S a m p l i n g a r e a 时间T i m e采样类群S a m p l i n g g r o u p 生态系统类型E c o s y s t e m t y p e PT M F *营养级传递趋势T r o p h i c t r a n s f e r t r e n d 备注N o t e s中国大鹿岛[46]D e e r I s l a n d ,C h i n a [46]2011.8双壳类㊁腹足类㊁海星㊁海葵B i v a l v e ,G a s t r o p o d ,A c t i n a r i a ,A s t e r o i d e a 海洋M a r i n e 0.2581.12生物放大趋势B i o m a g n i f i c a t i o n t r e n d 底栖食物链B e n t h i c f o o d c h a i n苏禄海[47]S u l u S e a [47]2002.11 12底层鱼类D e m e r s a l f i s h海洋M a r i n e <0.011.70生物放大B i o m a g n i f i c a t i o n波罗的海[57]B a l t i c S e a [57]1991 1993浮游植物㊁浮游动物㊁糠虾㊁鲱鱼P h y t o p l a n k t o n ,Z o o p l a n k t o n ,M y s i s a n d H e r r i n g海洋M a r i n e0.5881.01无明显趋势N o o b v i o u s t r e n d浮游食物链A p e l a g i c f o o d c h a i n意大利沿海区域[44]C o a s t a l a r e a ,I t a l y [44]2006.7沉积物㊁大型藻㊁海草㊁无脊椎动物㊁鱼S e d i m e n t ,M a c r o a l g a e ,S e a g r a s s e s ,I n v e r t e b r a t e s ,F i s h海洋M a r i n e<0.010.49生物减小B i o d i m i n u t i o n日本骏河湾[45]S u r u g a B a y ,J a p a n [45]2011.8,2012.9浮游动物㊁头足类㊁鱼类Z o o p l a n k t o n ,C e p h a l o p o d a n d F i s h海湾B a y<0.051.26生物放大B i o m a g n i f i c a t i o n北极巴芬湾[49]B a f f i n B a y [49]1998.3无脊椎动物㊁鱼类㊁鸟㊁哺乳类I n v e r t e b r a t e s ,F i s h ,B i r d a n d M a m m a l海湾B a y0.5071.03无明显趋势N o o b v i o u s t r e n d极地P o l a r r e g i o n南极阿德默勒尔蒂湾[50]A d m i r a l t y B a y ,A n t a r c t i c a[50]2012大型藻㊁端足类㊁海胆㊁腹足类㊁蛇尾㊁等足类㊁纽虫M a c r o a l g a e ,A m p h i p o d ,E c h i n o i d ,G a s t r o p o d ,O p h i u r o i d ,I s o p o d ,N e m e r t e a n海湾B a y N A #无明显趋势N o o b v i o u st r e n d极地,底栖食物链P o l a r r e g i o n ;b e n t h ic f o od c h a i n新喀里多尼亚珊瑚礁[48]C o r a l r e e f ,N e w C a l e d o n i a [48]2011藻类㊁无脊椎动物㊁鱼类A l g a e ,I n v e r t e b r a t e s a n d F i s h珊瑚礁C o r a l r e e fN A 1.11生物放大趋势B i o m a g n i f i c a t i o n t r e n d黄河河口[58]Y e l l o w R i v e r E s t u a r y ,C h i n a [58]2014.8无脊椎动物㊁鱼I n v e r t e b r a t e s a n d F i s h河口E s t u a r y 0.2641.12生物放大趋势B i o m a g n i f i c a t i o n t r e n d大辽河河口[59]D a l i a o R i v e rE s t u a r y ,C h i n a [59]2014.8无脊椎动物㊁鱼I n v e r t e b r a t e s a n d F i s h河口E s t u a r y 0.7910.98无明显趋势N o o b v i o u s t r e n d第1期杜森等:砷在海洋食物链中的生物放大潜力及发生机制探讨59㊀续表3研究区域S a m p l i n g a r e a 时间T i m e采样类群S a m p l i n g g r o u p生态系统类型E c o s y s t e m t y p e PT M F *营养级传递趋势T r o p h i c t r a n s f e r t r e n d 备注N o t e s孟加拉湾孙德尔本斯[60]S u n d a r b a n s m a n g r o v e e c o s y s t e m ,B a n g l a d e s h [60]2011.12植物㊁浮游动物㊁甲壳类㊁鱼P l a n t s ,Z o o p l a n k t o n ,C r u s t a c e a n s a n d F i s h红树林M a n g r o v e N A 无明显趋势N o o b v i o u s t r e n d 越南巴地头顿省[61]T r o p i c a l m a n g r o v e e c o s y s t e m ,V i e t n a m [61]2007.3悬浮颗粒物㊁头足类㊁甲壳类㊁鱼S u s p e n d e d p a r t i c u l a t e m a t t e r (S P M ),C e p h a l o p o d ,C r u s t a c e a n a n d F i s h红树林M a n g r o v e 0.5661.00无明显趋势/局部生物放大N o o b v i o u s t r e n d /p a r t l y b i o m a g n i f i c a t i o n 澳大利亚麦格理河口[51]L a k e M a c q u a r i e E s t u a r y ,A u s t r a l i a [51]2006.7碎屑㊁浮游动物㊁自养生物㊁食草动物㊁碎屑动物㊁食肉动物和杂食动物D e t r i t u s ,Z o o p l a n k t o n ,A u t o t r o p h s ,H e r b i v o r e s ,D e t r i t i v o r e s ,C a r n i v o r e s ,O m n i v o r e海草床S e a g r a s s N A局部生物放大P a r t l yb i o m a g n i f ic a t i o n阿根廷圣罗克水库[53]S a n R o q u e R e s e r v o i r ,A r g e n t i n a [53]2012.3浮游生物㊁虾㊁鱼P l a n k t o n ,S h r i m p a n d F i s h淡水F r e s h w a t e r <0.010.890生物减小B i o d i m i n u t i o n意大利马里内罗湖[55]T h e M a r i n e l l o p o n d s ,I t a l y [55]2009.5植物㊁无脊椎动物㊁鱼P l a n t s ,I n v e r t e b r a t e s ,F i s h淡水F r e s h w a t e r <0.010.779生物减小B i o d i m i n u t i o n阿根廷巴塔哥尼亚山湖泊[54]P a t a g o n i a m o u n t a i n l a k e ,A r g e n t i n a [54]2004 2009浮游生物㊁底栖生物㊁鱼㊁植物P l a n k t o n ,B e n t h i c o r g a n i s m s ,F o r a g e f i s h a n d P l a n t s淡水F r e s h w a t e r <0.010.602生物减小B i o d i m i n u t i o n加拿大耶洛奈夫[52]Y e l l o w k n i f e ,C a n a d a [52]2010.6浮游植物㊁浮游动物P l a n k t o n ,Z o o p l a n k t o n 淡水F r e s h w a t e r N A生物减小B i o d i m i n u t i o n浮游食物链P l a n k t o n i cf o o d c h a i n阿根廷科尔多瓦[56]C ór d o b a ,A r g e n t i n a [56]2015.8浮游植物㊁虾㊁鱼P l a n k t o n ,S h r i m p a n d F i s h淡水F r e s h w a t e r <0.010.851生物减小B i o d i m i n u t i o n注:#N A 表示无有效数据;T M F 表示营养级放大系数㊂N o t e s :#N A m e a n s n o v a l i d d a t a ;T M F s t a n d s f o r t r o p h i c m a g n i f i c a t i o n f a c t o r .60㊀生态毒理学报第14卷图2㊀砷沿食物链/网传递模式图注:(A)生物减小㊁(B)生物放大㊁(C)无显著趋势㊁(D)局部环节生物放大㊂Fig.2㊀The mode pattern of arsenic transfer along food chains/websNote:(A),biodiminution;(B),biomagnification;(C),no significant trend;(D),biomagnification in part of food chains.态系统中的食物链传递可以归纳为图2中的4种模式,即生物减小㊁生物放大㊁无显著趋势和局部环节生物放大㊂在海洋生态系统中,仅发现砷在意大利南部沿岸区域有显著生物减小的现象[44],研究人员对7种大型海藻㊁2种海草㊁5种无脊椎动物㊁9种鱼类和2种鸟类(羽毛)进行采样分析后发现,砷在沉积物㊁大型藻类和海草中的浓度最高(分别高达11.8㊁35.4和17.4μg㊃g-1),然后随着营养水平的增加而减少,在三级消费者和顶级捕食者(鱼类和鸟类)中的As浓度低于2μg㊃g-1,这些不同生物中As浓度的对数值(Log10As)与δ15N之间呈现出显著的负相关性,清晰地表明砷在该食物网中的生物减小作用㊂需要指出的是,这一研究区域严格上说并不是一个单一的食物链/网,而是一个开放区域的生物集合,因此其结果可能无法反映真实的砷食物链传递结果㊂相反,砷在其他的海洋生态系统中的食物链传递主要呈现出生物放大(图2B)㊁无显著趋势(图2C)和局部环节生物放大(图2D)等多种现象㊂在日本骏河湾生态系统中发现生物(浮游动物㊁头足类和鱼类)体内砷含量(Log10As)与营养级(δ15N值)呈显著正相关(P< 0.05),有着明确的生物放大作用[45]㊂砷在中国大鹿岛底栖食物链中也存在一定的生物放大趋势[46]㊂苏禄海底层鱼类体内砷含量(Log10As)与营养级(δ15N 值)之间同样存在显著的正相关(P<0.01),且BMF达1.70[47]㊂在珊瑚礁生态系统中,砷则出现了生物放大的趋势[48]㊂在南极和北极的极地海洋性生态系统中,砷的浓度与营养级之间无显著相关性[49-50]㊂在沿海的浅海生态系统(包括河口㊁红树林和盐沼海草床等),砷同样表现出了生物放大的潜能㊂Barwick等[51]对澳大利亚温带河口海草生态系统中食物网的研究发现,在碎屑㊁浮游动物㊁自养生物㊁食草动物㊁碎屑动物㊁食肉动物和杂食动物间的35条经验食物链中有19条出现砷浓度从被捕食者到捕食者增加的现象;在浮游动物(2.1μg㊃g-1)ң浮游双壳类动物(4.8μg㊃g-1ʃ0.9μg㊃g-1)ң肉食性鱼(6.2μg ㊃g-1ʃ0.4μg㊃g-1)和碎屑(6μg㊃g-1ʃ3μg㊃g-1)ң杂食性虾(8μg㊃g-1ʃ1μg㊃g-1)ң杂食性鱼(11μg㊃g-1ʃ2μg㊃g-1)2条食物链中观察到As的生物放大,而这2条食物链都属于底栖食物链㊂砷在淡水生态系统中的食物链传递研究相对较少,但是都一致地表现出显著的生物减小作用(图2A),仅在BMF值上有所差异[52-56]㊂例如,在阿根廷的一个湖泊调查中发现,砷在由浮游植物㊁底栖生第1期杜森等:砷在海洋食物链中的生物放大潜力及发生机制探讨61㊀物㊁鱼和植物组成的食物网中出现显著的生物减小现象(P <0.01)[54]㊂从这些研究结果中不难发现,砷沿海洋食物链/网传递具有一定的复杂性,但是普遍具有随营养级升高维持高生物累积或生物放大的潜力,与淡水食物链/网中砷的生物减小形成显著的对比㊂然而,这些研究均存在一定的局限性(例如采样没有覆盖主要生物类群,没有用δ15N 确定生物营养级,缺少对同一生态系统的重复性调查等),造成研究结果的诸多不确定性,使得砷在海洋食物链/网中是否生物放大仍然存在疑问㊂更重要的是这些研究基本都没有对不同砷形态进行分析,而砷形态的区别恰恰是海洋生物和淡水生物最大的差异,很可能是解析砷在海洋生态系统中生物放大潜力的关键因素㊂3㊀砷在海洋食物链/网中生物放大潜力的可能机制(Possible mechanisms of arsenic biomagnification potential along food chains /web )目前已有研究从生物动力学角度来分析无机砷的食物链传递,在几种海洋鱼类和无脊椎动物中测定了控制食物链传递的2个参数(同化率和排出率),通过模型计算表明,无机砷难以在食物链中被生物放大[18,35,37]㊂Zhang 等[62]研究了蛤和多毛类对沉积物中砷的生物动力学和生物可利用性,发现生物对砷的累积与沉积物中砷的形态有关,砷甜菜碱是有机砷中同化效率最高的形态,而As(III)的同化效率较低且排出速率较快㊂海洋鱼类对痕量元素的同化率受被捕食者亚细胞痕量元素的分布以及鱼类的摄食过程所影响[63],而不同的食物对海洋鱼类As 的生物有效性和生物累积存在显著的影响作用[64]㊂痕量元素能够在底栖食物链中生物放大的主要原因可能是腹足类能够有效吸收食物中的痕量元素,而浮游生物食物网生物减小的发生则可能是其对痕量元素较低的同化率和较高的排出率[65]㊂因此,砷的生物放大作用很可能是由有机砷的食物链传递作用所引起的㊂痕量元素的有机形态通常比无机形态具有更高的生物可利用性和食物链传递能力,其中了解最多的为汞㊂有机汞(以甲基汞为主)具有很强的亲脂性,可以直接通过细胞膜结构进入细胞并与生物分子结合,因此具有比无机汞更高的生物可利用性㊂而甲基汞的生物放大作用也远超无机汞,是痕量元素生物放大的最典型案例[7]㊂由此,有机砷可能比无机砷更容易被生物放大,这在一定程度上解释了砷在海洋和淡水生态系统中食物链传递的差异现象㊂在越南南部沿海的红树林生态系统中通过对悬浮颗粒物㊁头足类㊁甲壳类和鱼类的调查后发现,尽管总砷浓度(Log 10As)与营养级(δ15N 值)之间没有显著的相关性(P =0.556),但生物体内的砷甜菜碱出现了明显的生物放大现象(TMF =1.30,P <0.01)[61]㊂海洋鱼类和贝类普遍具有将从环境中吸收的无机砷通过生物转化合成砷甜菜碱的能力(如图3),这种生物转化过程包括As(V)还原为As(III),As(III)甲基化至一甲基砷和二甲基砷,以及经过后续未知的过程合成砷甜菜碱[66-68]㊂砷甜菜碱的合成,有助于海洋鱼类将砷累积在体内,从而达到较高富集的结果㊂在这种生理作用下,当鱼类长期暴露在高无机砷环境中时,导致的结果是较高砷甜菜碱的富集,这也是鱼类的一项重要解毒机制[69]㊂在温带海洋盐沼系统中,植物体内砷主要为无机砷,但摄食这些植物图3㊀海洋鱼类对砷的生物转化[67]Fig.3㊀Arsenic biotransformation of marine fish [67]62㊀生态毒理学报第14卷的动物体内砷的主要形态为砷甜菜碱和砷糖为主的有机砷,说明动物将吸收的无机砷生物转化成了有机砷[70]㊂Zhang等[37]通过构建了2条底栖食物链(沉积物 沙蚕Nereis succinea 诸氏鲻鰕虎鱼和沉积物 蛤Gafrarium tumidum 鰕虎鱼)研究了砷沿底栖食物链的传递和生物转化作用,发现蛤对砷的吸收效率分别为35%~65%,沙蚕对砷的吸收效率分别为52%~73%,从而造成沙蚕比蛤具备更高的砷累积能力;同时,在沉积物中砷的形态以无机砷为主,到初级消费者体内砷甜菜碱成为主要形态,再到鱼类体内砷甜菜碱可以占到95%以上,说明无机砷在底栖食物链中被高效转化,并主要发生在初级消费者环节,该结果也印证了As在底栖食物链中存在更大生物放大潜能㊂当砷从不同的饵料生物传递到海洋鱼类时,饵料中的有机砷比例与总砷的营养级放大系数(TMF)呈现显著的正相关,表明海洋生物中有机砷比无机砷具有更高的食物链传递能力,可以导致海洋鱼类富集更高浓度的砷[64]㊂Zhang等[67]研究了海洋鱼类(草食性篮子鱼Siganus fuscescens和肉食性鲈鱼Lateolabrax japonicus)对砷的营养转化和生物可利用性,发现食物中的无机砷较难被海洋鱼类吸收,而是在体内被生物转化成了有机砷,而食物中的砷甜菜碱可直接通过消化器官的上皮细胞被鱼类吸收,成为鱼体内砷的最终存储形态㊂对澳大利亚红树林的调查显示,红树林生态系统中的动物主要从食物中吸收有机砷(砷甜菜碱和砷核糖等)[71]㊂长期砷甜菜碱的暴露会导致大西洋鲑鱼(Salmo salar)和大西洋鳕鱼(Gadus morhua)肌肉中砷浓度显著增加[72]㊂目前关于有机砷食物链传递的生物动力学研究较少,因此需要对有机砷(特别是砷甜菜碱)的生物累积动力学参数进行量化解析,厘清其对总砷食物链/网传递的贡献㊂除了生物体中的砷形态(内因)可能直接作用于砷的食物链传递以外,海洋生态系统中的生物和环境因素(外因)也可能对其造成影响㊂环境因素如温度㊁浊度㊁营养盐和光照等可影响生物中的砷形态,间接作用于砷的食物链传递;也可能通过影响生物的生长速率和代谢率(特别是小个体生物),间接影响砷的摄入㊁代谢和排泄[73-74],这些作用可综合体现在砷生物累积的季节变化上㊂盐度能够影响大亚湾几种常见鱼类和甲壳类体内砷的累积,尤其是砷甜菜碱的浓度;同时沉积物中高砷的浓度对海洋鱼类体内砷的累积也有重要影响[75]㊂室内实验也证明高盐度会导致贻贝(Mytilus edulis)对砷甜菜碱的高累积[76]㊂有研究认为海藻通过细胞磷酸盐转运系统累积海水中的砷酸盐,但是其体内的砷糖水平更主要是受到海水中氮盐的影响[77]㊂食物网中生物组成的不同与所处环境的差异也会对砷的食物链传递产生影响㊂底栖无脊椎动物和鱼类往往比游泳生物含有更高浓度的砷,除了物种差异的原因外,还包括不同生物类群的食性不同,食物中砷浓度和形态的差异也会对砷的累积产生差异,因此,当食物网中生物组成偏向于底栖食性时,可能会导致更大的生物放大潜力[37,62]㊂因此,当研究砷的食物链传递时,不能仅着眼于砷本身而忽视了其他环境因素的作用㊂4㊀总结与展望(Summary and prospect)海洋生物普遍具有高含量的砷,这些砷主要为低毒性的有机砷形态㊂砷在淡水生态系统中普遍生物减小,但是在海洋生态系统中出现了一定的生物放大潜力㊂海洋鱼类和贝类普遍具有将从环境中吸收的无机砷通过生物转化合成砷甜菜碱的能力,有机砷比无机砷具有更高的食物链传递能力,可以导致海洋鱼类富集更高浓度的砷㊂因此,有机砷的存在可能有助于砷沿着海洋食物链/网富集,在某些情况下被生物放大㊂但是目前关于砷在海洋生态系统中生物放大潜力及机制的研究较为匮乏,今后应该从一下几个方向开展:(1)不同砷形态如何在海洋食物链/网中传递的研究㊂目前关于砷在生态系统中的传递主要是关于总砷的研究,缺乏对不同砷形态在传递的过程中累积㊁传递和转化的研究工作㊂(2)砷在海洋生态系统中生物放大潜力的影响因素㊂过去主要关注痕量元素在生态系统中传递的调查工作,应该系统地研究砷在不同海洋食物链/网中从初级生产者到高营养级生物中的行为,通过比较不同物种对砷累积的差异以及不同食物链之间的异同,厘清引起砷生物放大的关键因素㊂(3)砷甜菜碱合成关键基因及其调控机制的研究㊂海洋鱼类能累积较高含量的砷,其中的砷主要为砷甜菜碱,所以应该采用分子生物学等方法研究砷甜菜碱合成的关键基因和调控机制,阐明海洋生物高砷累积的调控机制㊂(4)室内模拟实验与野外调查相结合,通过不同砷形态食物链传递的室内实验来验证砷形态是否是导致海洋生态系统中砷放大的关键原因,同时验证。
海水 砷 标准物质

海水砷标准物质海水中的砷污染已经成为一个全球性的环境问题。
砷是一种有毒且具有慢性毒性的元素,对人类健康和生态系统的稳定性造成了不可忽视的影响。
为了保护环境和人类的健康,制定和遵守海水中砷的标准物质是至关重要的。
砷的存在形态和污染源砷在海水中以无机砷化物和有机砷化物的形式存在。
无机砷主要包括亚砷酸盐和砷酸盐,而有机砷通常以甲基砷酸盐和甲基砷酸二盐的形式存在。
这些不同形态的砷物质对人体和环境的影响程度各不相同。
海水中的砷主要来自于人类活动和自然过程。
人类活动如工业废水、矿区废水和农业排放等都会导致砷的污染。
此外,地壳物质的风化和火山活动也会释放出砷。
这些源头都对环境和人类健康构成了潜在威胁。
砷的危害和影响砷污染对环境和人类健康有着严重的危害和影响。
无机砷物质对人体健康具有慢性毒性,长期暴露可能导致癌症、皮肤疾病、心血管疾病和神经系统疾病等。
此外,砷还可以造成水生生物的死亡和生态系统的不稳定。
海水中的砷标准物质的制定意义制定和遵守海水中砷的标准物质对于保护环境和人类健康至关重要。
标准物质是为了控制和监测砷污染而制定的一套规定和方法。
它可以帮助政府和相关机构制定有效的控制措施,确保砷浓度在安全范围内。
海水中砷的标准物质制定过程制定海水中砷的标准物质需要经过科学研究和严格监测。
首先,科学家会收集海水样品,并对其中的砷含量进行分析。
接着,他们会评估砷对环境和人体的危害,并结合现有的科学研究和国际经验,制定适用于不同地区和用途的砷标准物质。
海水中砷的标准物质的具体内容海水中砷的标准物质通常包括以下几个方面:1. 砷的容许浓度:标准物质会明确规定海水中砷的容许浓度,以确保不会对人体健康和生态系统造成不可逆的损害。
这个容许浓度通常会根据不同的地区和用途进行细分。
2. 监测方法和频率:标准物质会详细规定监测砷的方法和频率,以确保数据的可靠性和一致性。
这些方法通常包括样品采集、样品处理、分析方法和数据统计等。
3. 管理措施:标准物质还会指导政府和相关机构采取合适的管理措施来减少砷的污染。
贝壳类海产品砷形态分析实验研究

贝壳类海产品砷形态分析实验研究王继霞;胡蔚娜;叶明德;张颜;陈帆【期刊名称】《实验技术与管理》【年(卷),期】2016(033)010【摘要】实验探讨了超声时间、温度、功率对贝壳类海产品中砷化合物提取的影响,对高效液相色谱-原子荧光(HPLC-AFS)联用技术测定贝壳类海产品中的 As 形态进行了实验研究.在优化的实验条件下,As(V)、一甲基砷(MMA)、二甲基砷(DMA)、砷甜菜碱(AsB)在5~200μg/L 范围内线性关系良好,相关系数均大于0.9995,4种砷化合物的检出限分别为1.47、3.34、3.30、2.36μg/L.样品加标回收率为92.3%~115.2%,相对标准偏差为1.04%~2.20%(n=6).研究结果表明,该方法操作简单、快速、灵敏度高,结果准确、可靠,适用于海产品中砷的形态分析.【总页数】5页(P45-48,54)【作者】王继霞;胡蔚娜;叶明德;张颜;陈帆【作者单位】温州大学化学与材料工程学院,浙江温州 325035;温州大学化学与材料工程学院,浙江温州 325035;温州大学化学与材料工程学院,浙江温州325035;温州大学化学与材料工程学院,浙江温州 325035;温州大学化学与材料工程学院,浙江温州 325035【正文语种】中文【中图分类】S944.3;O657.7【相关文献】1.甲壳类、贝壳类海产品气单胞菌分布与消长分析 [J], 张怡明;何水渊;罗学辉;张建群2.近海区域贝壳类海产品重金属含量探究 [J], 周丽群3.3种培养基对甲壳类、贝壳类海产品中气单胞菌检出情况分析 [J], 罗学辉;张怡明;何水渊;张建群4.超声辅助酶水解-高效液相色谱-氢化物发生-原子荧光光谱测定贝壳类海产品中砷形态 [J], 王继霞;张颜;叶明德;陈帆5.儿童缺锌应多吃贝壳类海产品 [J],因版权原因,仅展示原文概要,查看原文内容请购买。
海产品无机砷检测及含量控制技术研究

海产品无机砷检测及含量控制技术研究臧汝瑛;许振伟【摘要】现阶段在海产品无机砷检测及含量控制技术研究方面,国家海检局加大了检测监管和控制含量的力度,这是因为海产品中的无机砷过多,就会损害人的皮肤和诱发慢性肝脏病变,最终引起急性中毒或是致癌,给人类的身体健康和安全带来了极大的危害.目前我国海产品无机砷检测方法主要是原子荧光法,但这些技术还只是处于初步发展与研究的阶段,依然存在严重的不足,这就要求在今后的发展过程中,需要采用科学有效的无机砷检测及含量控制技术对海产品质量进行严格把关,以便能够提高海产品的质量安全和无机砷检测控制技术.【期刊名称】《食品工程》【年(卷),期】2014(000)001【总页数】3页(P41-43)【关键词】海产品;无机砷;检测;含量控制;原子荧光法【作者】臧汝瑛;许振伟【作者单位】威海市产品质量监督检验所,山东威海264209;威海市产品质量监督检验所,山东威海264209【正文语种】中文【中图分类】TS207.5+1现阶段由于工业的快速发展,使得许多地下水源都被严重污染,这些被污染的水源流入海中就会使大量的海产品富营养化,从而让海产品成为含砷量较高的食物。
这些海产品中的砷元素大多数为无机砷形式,一旦长时间积累就会给人的身体带来巨大的危害。
面对这种危害,对海产品无机砷检测及含量控制技术进行深入的分析和研究就显得十分重要了。
一方面通过无机砷检测可以大大提高我国海产品的质量,很好的保障了消费者的人身安全,同时也可以提高我国海产品无机砷检测及含量控制技术的研究水平。
另一方面,通过对海产品进行无机砷脱离处理,也可以使我国的海洋生物资源得到充分的利用,对我国海洋资源的开发是很有利的。
尽管现阶段有许多的无机砷检测方法(AAS、AES、HPLC等技术),但是常用的而且比较成熟的方法(原子荧光标记检测法)还是较少,远远不能满足整个海产品无机砷检测及含量控制的要求。
现今,几乎所有的海产品养殖和捕捞都是在海水中来进行的,而海水中的无机砷经过长期的积累,使得大多数海产品中都含有无机砷化合物(主要包括五价砷和三价砷),许多海产品的砷元素含量甚至都超过了我国法定卫生元素含量标准。
砷对植物生长的影响及抗氧化系统响应机制研究进展

砷对植物生长的影响及抗氧化系统响应机制研究进展徐鼎;刘艳丽;杜克兵;王晓湘;王滑;涂炳坤【摘要】砷污染越来越受人们的关注,已成为迫切需要解决的环境污染问题之一.本文分析了砷对植物生理活动低促高抑的影响及植物在不同砷价态下的生理响应;阐述了As(Ⅲ)、As(V)在植物体内的吸收、转运和外排机理,着重探讨植物体内抗氧化酶类、植物螯合肽(PC)、谷胱甘肽(GSH)等对砷的响应机制,为植物修复砷污染土壤的研究提供参考.【期刊名称】《湖北林业科技》【年(卷),期】2014(043)001【总页数】8页(P8-15)【关键词】砷;植物;生长影响;抗氧化系统;解毒;耐性机理【作者】徐鼎;刘艳丽;杜克兵;王晓湘;王滑;涂炳坤【作者单位】华中农业大学园艺林学学院武汉 430070;华中农业大学园艺林学学院武汉 430070;华中农业大学园艺林学学院武汉 430070;华中农业大学园艺林学学院武汉 430070;华中农业大学园艺林学学院武汉 430070;华中农业大学园艺林学学院武汉 430070【正文语种】中文【中图分类】X59随着工业发展和农业生产的现代化,重金属污染已经成为危害全球环境质量的一个重要问题。
砷污染在国内外作为一个非常严峻的环境问题,已经受到人们的普遍关注。
最近的调查发现,非洲布基纳法索的雅滕加省的一个村庄有240人患有皮肤病,其中29.26%和46.34%的被确诊为黑变病和角化病,这与当地的井水中含有过量(超过世界卫生组织的规定值10 μg·L-1)的砷有关[1]。
1998年报道过印度、孟加拉、智利等20余个国家存在饮水砷中毒的现象[2]。
目前,全球至少有5 000多万人口仍然面临着地方性砷中毒的威胁,其中大多数为亚洲国家,然而中国也是受砷中毒最为严重的国家之一,包括内蒙、新疆、陕西、湖南、云南、贵州等省区受到砷污染的影响均比较严重。
砷污染的来源极其广泛,环境中砷污染主要是人类的工农业生产活动造成的。
海带含砷问题的探讨

海带含砷问题的探讨
孙建璋;孙海藻
【期刊名称】《海洋水产科技》
【年(卷),期】1999(000)001
【摘要】本文从海带含砷量及其化学成分,药理试验,国际及国内食品卫生有关规定等方面进行分析讨论,从食品卫生学角度阐明海带中砷毒性很小,食有海带有益人类健康。
【总页数】4页(P38-41)
【作者】孙建璋;孙海藻
【作者单位】浙江省苍南县水产研究所;浙江省苍南县水产研究所
【正文语种】中文
【中图分类】TS253.58
【相关文献】
1.含砷物料资源化利用与无害化处置技术路线探讨 [J], 曲胜利;董准勤;陈涛;曲超
2.含砷硫精矿制酸问题的探讨 [J], 吴桂荣;李伟达
3.海带(Laminaria japonica Aresch)含砷问题的探讨 [J], 孙建璋;孙庆海
4.焦锑酸钠生产中含砷用液治理方法的探讨 [J], 周贝娜
5.焦锑酸钠生产中含砷用液治理方法的探讨 [J], 周贝娜
因版权原因,仅展示原文概要,查看原文内容请购买。
新时期海产品无机砷检测与含量控制策略

新时期海产品无机砷检测与含量控制策略
孙枫林;鞠光秀;赵雅婷
【期刊名称】《食品界》
【年(卷),期】2022()10
【摘要】海产品无机砷检测与含量控制技术的应用,对提高海产品无机砷检测监管与含量控制水平等有促进作用。
由于海产品中的无机砷过多,对人体身体健康安全等会产生直接的影响。
因此,从技术分析的角度,对海产品无机砷检测与含量控制进行分析,则需要对无机砷的含量进行检测与分析,并对海产品无机砷含量进行控制,旨在提高海产品无机砷含量检测可靠性,并对其进行有效控制,提高海产品的安全性。
【总页数】3页(P111-113)
【作者】孙枫林;鞠光秀;赵雅婷
【作者单位】青岛市疾病预防控制中心;胶州市疾病预防控制中心
【正文语种】中文
【中图分类】R15
【相关文献】
1.余姚市海产品无机砷含量抽样检测分析
2.海产品无机砷检测及含量控制技术研究
3.舟山加工海产品中无机砷含量分析及评价
4.海产品无机砷检测及含量控制技术分析
5.不同产地保健食品原料中总砷及无机砷含量检测
因版权原因,仅展示原文概要,查看原文内容请购买。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
砷存在的形态[13~16]和海产品中各种形态砷化物的含
集,特别是以水体中浮游生物为食物链的水产生物体 量。
内有明显的富集倾向,致使海产品中有较多的砷[12]。
表 1 海产品中砷的存在的形态
Table 1 Arsenic species in seafood
名称
存在
化学分子式
亚砷酸盐(As(III))
砷为有毒有害元素,可从人畜的消化道和呼吸道 血性腹沟。重者因高度脱水,引起休克、少尿、蛋白
257现代食品科技尿以 Nhomakorabea昏睡、痉挛[29]。
Modern Food Science and Technology
As(V)
2014, Vol.30, No.11 2.1
表 2 海产品中各种形态的砷化物含量(mg/kg 湿重)
有机砷是海产品中砷存在的主要形态。在鱼类及 进入体内,被胃肠道和肺脏所吸收,并散布于身体组
其他海产品中,超过 90%的砷是以毒性较小的有机形 织和体液中。所以,水体、食物和大气的砷污染,都
态存在的,无机砷的含量仅占 0.02%~11%[23~25]。海产
会直接危及人畜的健康。在体内,砷可与细胞内巯基
二甲基砷酸(DMA)
绿藻:海松
(CH3)2AsO(OH)
二甲基砷氧化醋酸(DMAA) 鳕鱼,扇贝,褐藻
(CH3)2As(O)CH2COOH
二甲基砷氧化乙醇(DMAE) 鳕鱼,扇贝,褐藻
(CH3)2As(O)CH2CH2OH
二甲基砷氧化丙酯(DMAP) 鳕鱼,扇贝,褐藻
(CH3)2As(O)CH2CH2COOH
(As(V))、砷胆碱(AsC)、砷甜菜碱(AsB)、砷糖、砷脂等。各种形态的砷通过不同代谢途径产生不同的代谢产物,从而表现出不
同的毒性。砷的形态主要是通过高效液相色谱与电感耦合等离子体质谱仪的联用(HPLC-ICP-MS)来测定的。有机砷中砷胆碱(AsC)
的代谢产物为砷甜菜碱(AsB)和砷脂;砷甜菜碱(AsB)经尿液直接排出;不同的砷脂代谢产物不一样;砷糖的代谢产物至少 12
Mechanisms
WANG Ying, CHEN Miao-miao, TAN Ting-ting, LI Yi, TENG Jiu-wei (Department of Food Science and Engineering, College of Science and Engineering, Jinan University, Guangzhou 510632,
品中砷的总量,受海产品种类及所在地区影响。中国 酶结合而使其失去活性,从而影响组织的新陈代谢,
常用的海产品里水藻中的总砷浓度最大[2];挪威各海 引起细胞死亡;也可使神经细胞代谢障碍,造成神经
产品里鳕鱼中砷的平均值最高[18];地中海不同区域里
系统病变;对于消化道,砷有腐蚀性,接触部位可产
鳕鱼中砷的含量最高,鲭鱼中砷的含量最低[26]。海产
现代食品科技
Modern Food Science and Technology
2014, Vol.30, No.11
海产品中的砷及其代谢机制的研究进展
王瑛,陈苗苗,谭婷婷,李祎,滕久委
(暨南大学理工学院,广东广州 510632)
摘要:海产品中的砷通过食物链富集的方式进入人体。海产品中检测出的砷形态有三价的亚砷酸盐(As(III))、五价的砷酸盐
Key words: arsenic; seafood; speciation; toxicity; cooking; detection; metabolism
海产品营养丰富,味道鲜美,并且具有低脂肪、 高蛋白的特点,深受人们的喜爱。海产品中含有大量 不同形态的砷[1~3]。在美国人的饮食中,90%的砷来源 于海产鱼[4]。而在加拿大和日本,也有 70%的砷来源 于海产品[5~6]。人们在食用含砷的海产品时,由于砷在 人体内易发生富集,对人体的生命健康会造成一定威
As(III)+MMA <LOD
Table 2 Concentrations of arsenic species in seafood (mg/kg wet
DMA
0.6
weight)
AsB
5.4
总类
国家
砷形态
含量 参考文献
AsC
<LOD
总砷
1220
[16]
总砷
0.45
[2]
DMAA
0.013
挪威
DMAP
生急性炎症、出血与坏死;砷进入肠道,可引起腹泻,
品中砷的含量与其所在水环境以及饮食习惯有很大关 并可使心脏及脑组织缺血,引起虚脱,意识消失及痉
系。生活在海洋底部的鱼(如比目鱼)所含砷的浓度 挛等。砷被吸收后,可麻痹血管运动中枢,直接作用
较高;生活在海洋地表的贝类也有较高浓度的砷[27]; 于毛细血管,使脏器的微血管发生麻痹、扩张和充血,
<LOD
砷糖
中国
DMA
0.02 0.029
褐藻
DMAE
0.005
海藻,羊栖菜
As(OH)3
砷酸盐(As(V)) 砷甜菜碱(AsB) 砷胆碱(AsC)
海藻,羊栖菜 鱼,节足动物,软体动物
虾类
AsO(OH)3 (CH3)3As+CH2COO(CH3)3As+CH2CH2OH
三甲基氧化砷(TMAO) 四甲基砷离子(TMAs+)
鱼,海螺 贝类,海葵等
(CH3)3AsO (CH3)4As+
China)
Abstract: Arsenic in seafood enters the human body by a process of food chain enrichment. The arsenic species detected in seafood include arsenide (As(III)), arsenate (As(V)), arsenocholine (AsC), arsenobetaine (AsB), arsenosugars, and arsenolipids. Arsenic species are metabolized through various pathways and result in different metabolites and toxicities. High-performance liquid chromatography and inductively coupled plasma mass spectrometry (HPLC-ICP-MS) was used to analyze arsenic species in seafood. The metabolites of AsC, a type of organic arsenic, were mainly AsB and arsenolipids. AsB could be excreted through urine. Each arsenolipid has different metabolites. More than 12 arsenic species were found as metabolites of arsenosugars, of which dimethylarsinic acid (DMA(V)) was the most common. The pentavalent methylated DMA(V), and trivalent methylated DMA(III) were the final metabolites of inorganic arsenic species. Cooking methods affect the concentration of total arsenic and the form of arsenic. Some methods can result in increased toxicity. Therefore, to prevent excessive intake of toxic forms of arsenic, effective measures including reducing seafood intake, increasing intervals between seafood meals, daily intake of specific nutrients, and choosing optimal cooking methods, should be recommended to consumers.
种,最主要的是五价的二甲基砷酸盐(DMA(V))。无机砷(包括砷酸盐和亚砷酸盐)最后代谢产物为五价的二甲基砷酸盐(DMA(V))
和三价的二甲基亚砷酸盐(DMA(III))。海产品经过不同烹调处理后,其总砷浓度或多或少都有增加,砷形态也有变化,部分加工会
导致毒性增加。对于消费者来说,注意饮食习惯包括海产品摄入量,摄入间隔,日常营养元素摄入与烹调方式的选择等是预防过量摄
估模型。
(DMAA)、二甲基砷氧化丙酯(DMAP)、三甲基砷
1 海产品中砷的形态
氧化丙酯(TMAP)、三甲基砷氧(TMAO)、四甲基 砷离子(TMAs+)、砷胆碱(AsC)、砷甜菜碱(AsB)、
环境中的砷污染物经过雨水冲刷地表径流等各种 砷糖、以及砷脂等。表 1 和表 2 分别列举了海产品中
途径进入到海水中,又通过食物链在海产生物体内富
长期生活在砷浓度较高的海水中的鱼摄入了高浓度的 以致血压下降;并且被人体吸收的砷,部分贮留于肝
砷后,未代谢完的砷在体内聚集,而使其所含的砷比 脏,会引起肝细胞退行形变和糖原消失[28]。
长期生活在新鲜的湖水中的鱼所含的砷浓度高。
急性砷中毒的病人往往出现全身不适、疲乏、无