分子生物学第七章教程文件
分子生物学 第七章 蛋白质的生物合成

7.1 遗传密码 7.2 tRNA 7.3 氨酰-tRNA合成酶 7.4 核糖体 7.5 与蛋白质合成有关的因子 7.6 蛋白质合成的生物学机制 7.7 蛋白质转运机制
比DNA复制和转录更为复杂的过程
氨基酸活化与转运----这个过程是在氨基酸活化酶和镁 离子作用下把氨基酸激活成为活化氨基酸。 起始----核糖体与mRNA结合并与氨酰基tRNA生成起 始复合物。 延伸----由于核糖体沿mRNA5’端向3’端移动,开始了 从N端向C端的多肽合成,这是蛋白质合成过程中速度 最快的阶段。 终止以及肽链的释放,核糖体从mRNA上解离,准备 新一轮合成反应。
(3)校正tRNA 校正tRNA分为无义突变及错义突变校正。 无义突变:在蛋白质的结构基因中,一个核 苷酸的改变可能使代表某个氨基酸的密码 子变成终止密码子(UAG、UGA、 UAA),使蛋白质合成提前终止,合成无 功能的或无意义的多肽。
错义突变是由于结构基因中某个核苷酸
的变化使一种氨基酸的密码变成另一种 氨基酸的密码。 错义突变的校正tRNA通过反密码子区的 改变把正确的氨基酸加到肽链上,合成 正常的蛋白质。(摆动原理现象)
结合的AA-tRNA分开。
7.1.3 遗传密码的特点
⑴连续性
⑵简并性与偏爱性
⑶通用性与专一性
(特殊性)
⑷终止密码
⑸密码子与反密码子
简并性: 由一种以上密码子编码同一个氨基酸的现象。 18种氨基酸都有一个以上的密码子 。
同义密码子。
只有精氨酸是个
例外,因为在真 核生物中CG双联
7.2 tRNA
7.2.1 tRNA的结构 由于链内的碱基互 补配对,tRNA呈现
分子生物学第七章原核生物基因表达调控

原核生物基因表达调控的特点
01
原核生物基因表达调控通常由特 定的转录因子、RNA聚合酶以及 其他调控蛋白介导,通过与DNA 的结合或解离来调节基因转录。
02
原核生物基因表达调控具有快速 响应环境变化的特点,能够在短 时间内调整基因表达模式,以适 应外界刺激和压力。
翻译后加工的调控
翻译后加工的调控
在翻译后加工阶段,新合成的蛋白质经过一系列修饰和加工,最终成为具有生物学活性的蛋白质。原 核生物通过控制翻译后加工酶的合成和活性来调控翻译后加工过程。此外,原核生物还可以通过控制 蛋白质的稳定性来影响其功能和表达水平。
总结
翻译后加工是基因表达调控的重要环节,原核生物通过控制翻译后加工酶的合成和活性,以及蛋白质 的稳定性来精细调控基因表达。
翻译延伸的调控
翻译延伸的调控
在翻译延伸阶段,核糖体沿着mRNA移动,将氨基酸组装成蛋白质。原核生物通过控制翻译延伸因子的合成和活 性,以及核糖体的合成和组装来调控翻译延伸。此外,原核生物还可以通过控制mRNA的结构和稳定性来影响翻 译延伸。
总结
翻译延伸是基因表达调控的重要环节,原核生物通过控制翻译延伸因子的合成和活性,以及核糖体的合成和组装, 以及mRNA的结构和稳定性来精细调控基因表达。
翻译起始的调控
原核生物通过控制翻译起始来调控基因表达。在翻译起始阶段, mRNA与核糖体结合,招募翻译所需的起始因子和其他成分。原 核生物通过控制起始因子的合成和活性,以及mRNA与核糖体的 结合来调控翻译起始。
总结
翻译起始是基因表达调控的重要环节,原核生物通过控制翻译起 始因子的合成和活性,以及mRNA与核糖体的结合来精细调控基 因表达。
92184-分子生物学-第七章 蛋白质变异效应与短肽

线 粒 体 呼 吸 链 膜 蛋 白 复 合 体 Ⅱ 的 三 维 结 构
HIV-1亚型膜蛋白V3环的氨基酸变异引起的效应
不同HIV-1亚型膜蛋白V3环的氨基酸序列特征及变异 特点,病毒V3顶端的四肽特征主要为: AE亚型GPGQ 48% 、B亚型GPGR 36%、其他形式16% 。
还发现DQDR、DQGQ等少见V3环顶端四肽组成形 式。
当tau蛋白发生高度磷酸化、异常糖基化、异常截 断作用以及泛素蛋白化时,tau蛋白就失去对微管 的稳定作用,神经纤维退化,功能丧失,引起中 枢神经系统变性疾病。
第三节 错误折叠蛋白质引起的变异效应
错误折叠蛋白质通常产生三种效应:
被细胞防御机制降解,导致功能缺失; 发生错误定位,导致细胞功能紊乱; 错误折叠蛋白质相互聚集形成淀粉样沉积
线粒体膜蛋白氨基酸突变引起一些疾病
人类有很多疾病诸如嗜铬细胞瘤、副神经节瘤和李氏症等 均与线粒体复合物II的翻译提前终止或氨基酸突变相关, 这些疾病多表现为氧自由基引起的神经性紊乱,而氧自由 基的产生与电子在线粒体复合物II中传递中的泄漏有关。
通过线粒体复合物II的结构,对已知的与人类疾病相关的 该复合物的突变位点进行了精确定位,发现这些突变位点 均位于电子传递体或醌结合位点的周围,它们的突变或影 响了电子传递体的结合,或影响了醌的结合,从而导致电 子传递的中断,逃逸到线粒体基质中或线粒体内膜中,产 生大量氧自由基进而导致肿瘤。
研究发现穿膜肽可以双向穿越细胞膜。它还可以介导如 DNA、多胺类,甚至比自身分子量大很多倍的寡聚核苷酸 进入细胞内。值得一提的是,穿膜时,分子是作为一个整 体插入,而不需要解折叠。该分子完全可以作为一种全新 的载体,它不需要能量,可以挂载的分子的类型多,分子 量大。
医学分子生物学 7 分子生物学常用技术

5′
3′
*********G —OH
*********C T T A A — P
3′
5′
5′
3′
P A A T T C*********
OH— G*********
3′ 5′
EcoR I: dATP+ [α-32P]-dTTP
19
根据反应原理确定同位素反应底物名称
• DNA的切口平移标记法 • 随机引物标记法 • DNA的3′末端标记
• 极高的特异性
缺点 半寿期短, 故要随用随标 放射性污染
9
1. DNA探针放射性标记方法
(1) 切口平移标记法 (2) 随机引物标记法 (3) PCR标记法 (4) 5′-末端标记 (5) 3′-末端标记
10
(1) DNA的切口平移标记法
5′
3 ′ 双链DNA
3′
5′
DNase I,Mg2+
② 反应产物的长度与加入的寡核苷酸引物的量呈反比。
当需要较长片段探针,可适当减少随机引物的加入量。
③ 所得到的标记产物为新合成的DNA单链。当采用单链
DNA片段或RNA作为模板时,必须注意得到的标记探针并 不是其本身.
④ 反应条件要控制在pH值6.6,抑制Klenow DNA聚合酶
的3′-5′外切酶的活性。
第七章 分子生物学常用技术
1
第一节 核酸分子杂交
2
*核酸分子杂交:
具有一定互补序列的不同来源的核苷酸单 链在一定条件下按照碱基配对的原则形成
杂交双链的过程。
实质: 核酸分子的变性与复性过程
变性:将双链DNA分子解聚成为单链的过程 复性:使单链聚合成双链的过程,又称为退火. 特点:高度特异性和灵敏性 杂交的双方: (靶,target) --待测序列
分子生物学 第7章 RNA复制与逆转录ppt课件

精选ppt课件2021
19
tRNA
R U5 PB
gag
R′ U′5
R U5 PB
gag
R′ U′5
PB
gag
负链 DNA
An
pol
env
U3 R
以 tRNA 为引物开始合成 cDNA 负链
pol
env
U3 R An
RNase H 降解模板 R 和 U5
An
pol
env
U3 R
负链 DNA3′端与模板 R 区配对(第一次跳跃)
精选ppt课件2021
11
7.2.1 RNA复制酶类
由病毒自己编码的RNA复制酶主要是依赖RNA的RNA 聚合酶(RNA-dependent RNA polymerase,RdRP)。
RNA复制酶功能与特性: ⑴ 以“–RNA”为模板合成“+RNA”活性高,而以“+RNA” 为模板合成“–RNA”活性低; ⑵ 合成RNA链方向为5′→3′; ⑶ 能将互补的双链RNA分开; ⑷ 缺少核酸酶活性,校正功能低,差错率高; ⑸ 多数分布在细胞质中,少数在细胞核中; ⑹ 酶活性具有可调控性,复制时,RdRP抑制外壳蛋白表达, 而成熟期,外壳蛋白抑制RdRP活性。
这类病毒包括滤泡性口腔炎病毒、流感病毒、
副流感病毒、莴苣坏死黄化病毒、麻疹病毒、狂犬病毒
等。
脱壳
病毒
RdRP
加工
亲代 –RNA
+RNA ① RdRP
子代
mRNA ② 翻译
–RNA
GpppN GpppN
帽子 mRNA
③ 翻译
子代病毒颗粒 组装
RNA 复制酶 病毒外壳蛋白 致病蛋白
分子生物学(全套课件557P)

分子生物学(全套课件557P)简介分子生物学是研究生物分子结构、功能和相互作用的学科。
它涉及到核酸、蛋白质和其他生物分子的研究,以及它们在细胞和生物体中的功能。
本文档是一套全面的分子生物学课件,共有557页。
本课件旨在帮助读者系统地了解分子生物学的各个方面,包括基本的分子生物学原理、实验技术、研究方法以及应用等。
目录1.第一章:分子生物学概述2.第二章:DNA结构与功能3.第三章:RNA结构与功能4.第四章:蛋白质结构与功能5.第五章:基因表达调控6.第六章:基因突变与遗传变异7.第七章:分子生物学实验技术8.第八章:分子生物学研究方法9.第九章:分子生物学的应用领域第一章:分子生物学概述1.1 什么是分子生物学分子生物学是研究生物体内分子的结构、功能以及相互作用的学科。
它涉及到DNA、RNA、蛋白质等生物分子的研究,以及它们在细胞和生物体中的功能。
1.2 分子生物学的历史与发展分子生物学起源于20世纪50年代,当时发现DNA是物质遗传信息的携带者后,科学家们开始研究DNA的结构和功能,从而奠定了现代分子生物学的基础。
1.3 分子生物学的重要性分子生物学的研究对于了解生命的本质和机理至关重要。
它不仅有助于解释遗传现象,还可以揭示细胞的结构、功能和调控机制,甚至为疾病的诊断和治疗提供理论基础。
2.1 DNA的组成与结构DNA是由基因序列组成的生物分子,它由核苷酸组成。
本节将介绍DNA的基本结构、双螺旋结构和碱基对的配对方式。
2.2 DNA复制与遗传信息传递DNA复制是细胞分裂过程中最重要的事件之一,它确保了遗传信息的传递和稳定性。
本节将介绍DNA复制的过程和机制。
2.3 DNA修复与突变DNA在生物体内容易受到各种外界因素的损伤,因此细胞拥有多种修复机制来修复DNA损伤。
本节将介绍DNA修复的方式和维护基因组稳定性的重要性。
3.1 RNA的种类与功能RNA是DNA转录的产物,它在细胞内发挥着多种功能,包括mRNA的编码信息传递、tRNA的氨基酸运载和rRNA的构建核糖体等。
分子生物学第7章RNA加工和核糖核蛋白
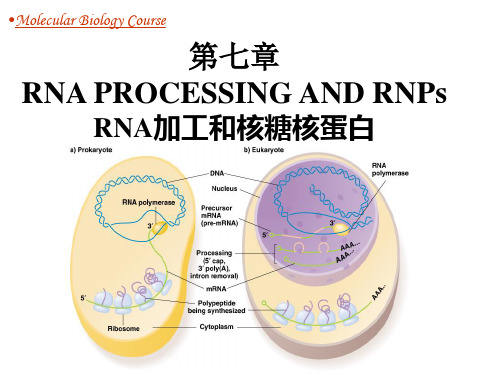
每个转录单位转录 成单个的RNA前体 分子,经剪切后变 成为成熟的RNA的 分子。
rRNA前体的加工 是由RNase Ⅲ负责 的。
2. 大肠杆菌rRNA转录后加工
➢ 6500nt初始转录物折叠形成一 些茎环结构
➢ 初始转录物与蛋白质结合形成 核糖核蛋白复合体
➢ 进行甲基化修饰 S-adenosylmethionine (SAM, S-腺苷甲硫氨酸)
RNA加工的类型
RNA加工的类型 ➢内切核酸酶和外切核酸酶切除核苷酸 ➢在初始转录物或剪切产物的5’端或3’端加上 核苷酸 ➢对某些核苷酸的碱基或糖苷进行修饰 ➢RNA编辑
第1节 rRNA加工与核糖体
一、原核生物rRNA 加工 二、真核生物 rRNA 加工
一、原核生物rRNA 加工
1. rRNA 基因的结构 ➢ E.coli 有7种不同的rRNA操纵子分散在整个基因组 中 ➢ 每个操纵子包含一个拷贝的5s rRNA、16s rRNA、 23s rRNA序列。 ➢ 有一到4个编码tRNA的序列。
(4) 最后修正:通过外切酶等将20S中和已退火的32S 中残余的ITS切除掉。
47S前 rRNA经历了一系列剪切,先切去外部转录间隔区,再切去内部 转录间隔区,释放32S和20S两个前RNA,最后释放18S、5.8S、28S rRNA。
47S pre-rRNA
ETS:external transcribed spacers ITS:internal transcribed spacers
rplJ(L10) rplJ(L10) rpoB(β亚基) rpoC (β’亚基)
转录
前体多顺反子mRNA
RNase Ⅲ 加工
成熟的mRNA
T7噬菌体早期区转录单条前体RNA,经 RNase Ⅲ剪切成5条成熟的mRNA
分子生物学第7章 DNA的重组与转座

Barbara McClintock 芭芭拉·麦克林托克) (芭芭拉 麦克林托克) 19021902-1992
C基因-Ds基因-Ac基因: Ac-Ds 控制系统 基因-Ds基因-Ac基因: Ac基因 基因 《玉米易突变位点的由来与行为》(1950),《染色体结构和基 玉米易突变位点的由来与行为》(1950), 因表达》 因表达》(1951)
7.2.2 细菌的特异位点重组
鼠伤寒沙门氏菌H1鞭毛蛋白和 鞭毛蛋 鼠伤寒沙门氏菌 鞭毛蛋白和H2鞭毛蛋 鞭毛蛋白和 白抗原表达的互变,称为鞭毛相转变, 白抗原表达的互变,称为鞭毛相转变, 这种转变是由H片段倒位决定的 片段倒位决定的, 这种转变是由 片段倒位决定的,此倒 位是由于特异重组位点hix发生重组后引 位是由于特异重组位点 发生重组后引 起的。 起的。
整合位点---attB、attP ◘ 整合位点 切除位点---attL、attR 切除位点 整合过程需要λ整合酶 ◘ 整合过程需要 整合酶 (integrase Int)( 编码) )(λ编码) )( 编码 和寄主的整合宿主因子IHF 和寄主的整合宿主因子 (integration host factor) ) 共同作用 ◘ 整合后的附着位点为 attL(BOP’) ( ) attR(POB’) ( )
Ⅰ型:两末端有相同的IS序列,或正向,或反向。 两末端有相同的IS序列,或正向,或反向。 IS序列 Ⅱ型:末端有反向重复序列。如TnA家族 末端有反向重复序列。 TnA家族
Tn A家族: A家族: 家族
TnA是复制型转座的转座子,长约5kb左右 TnA是复制型转座的转座子,长约5kb左右 。 是复制型转座的转座子 5kb 两端具有末端反向重复序列(而不是IS),长约38bp左右 IS),长约38bp左右, 两端具有末端反向重复序列(而不是IS),长约38bp左右, 任一个缺失都会阻止转座。 任一个缺失都会阻止转座。 中部的编码区编码三个基因:转座酶,解离酶和抗性基因, 中部的编码区编码三个基因:转座酶,解离酶和抗性基因, TnA家族都带有抗性标记 TnA家族都带有抗性标记 。 靶位点具有5bp的正向重复序列。 5bp的正向重复序列 靶位点具有5bp的正向重复序列。 解离位点( TnA家族特有的内部位点 家族特有的内部位点。 解离位点(res)是TnA家族特有的内部位点。
分子生物学课件第七章 RNA的生物合成

(for emergencies)
E. coli also has six alternative sigma factors that are used in special circumstances
Some of the sigma factors found in Bacillus subtilis
Control of sporulation in Bacillus subtilis also involves antisigma factors and anti-anti-sigma factors!
真核生物的启动子
三种RNA pol; 不同的RNA pol识别不同的启动子;
Promoter of RNAP II
原核生物RNA聚合酶可以被 利福霉素(rifamycin) 抑制。
利福霉素是从 链霉菌(streptomyces) 中分离得到的一 种抗生素,可以与细菌RNA pol的β亚基直接结合,抑制第 一个磷酸二酯键的形成。
人工合成的利福平 rifampicin,可供口服,用于细菌感染 的临床治疗。
真核生物叶绿体RNA聚合酶和线粒体RNA聚合酶对 -鹅膏蕈碱不敏感,但可以被利福霉素抑制。
RPB5 RPB6 RPB7
RPB8 RPB9 RPB10 RPB11 RPB12
220 Binds DNA
’
(and has CTD)
150 Binds NTPs
45
assembly
1
32
Not essential,
(stress response)
27
Target for activators
23
Efficient assembly
1分子生物学第七章基因作图和基因文库

原核生物重叠基因 • a. 一个基因完全在另一个基因里面。 • b. 部分重叠。 • c. 二个基因只有一个碱基对的重叠。 • 现在还不知道重叠基因在所有生物中是否具
有普遍性,仅在个别生物中发现有此现象, 例如流感病毒、大肠杆菌噬菌体G4,Qβ等
• 重叠基因的现象反映了原核生物利用有限的 遗传物质表达更多生物功能的能力。
• 一些物种之间的复杂性变化范围并不大,但 是C值却有很大的变化范围
• 这些问题的解决有待于进一步的研究。
基因组的基因数目
• 由于DNA中存在非编码序列,使计算产生 误差,故须先确定基因密度。
• 如果已知基因组的DNA序列,就可以获得 基因密度,包括ORF的平均长度和ORF间的 平均距离。
• 再根据已知基因数目及与已知基因相关的基 因数目来推测未知基因数目
• 虽然交换是一个普遍存在的遗传现象,但是,由 于当时还从来没有发现过交换发生在一个基因内 部的现象,因此在很长的一段时间里(直到40年 代中期)认为基因是三合一体,即既是一个功能 单位,也是一个突变单位和一个交换单位。
• (4)1944年Avery等首次用实验证实基因是 由DNA构成的。
• 基因是DNA
人
3.3×109 65000~8
0000
•真菌 •支原体(没有真正的细胞壁,格兰 氏阴性并且生长时需要类固醇)
•乳头(状瘤)多瘤空泡(形)病毒
•牛痘病毒
•细小病毒(小脱氧核糖核酸病毒)
Figure 1.36
The amount of
• 结构和功能越复杂,需要的基因产物的种类越 多,也就是说需要的基因越多,因而C值越大?
• 但是生物体复杂性和DNA含量之间并非线性关系(即 C值矛盾的本质)
分子生物学第七章原核生物基因表达调控

(三)、阻遏物 lac I 基因产物及功能
Lac 操纵子阻遏物 mRNA 是由弱启动子控制下组 成型合成的,该阻遏蛋白具有4个相同的亚基,每个亚 基均含347个氨基酸残基。
lacI 基因为组成型,通过启动子的上升突变体可获 得较多的阻遏蛋白;
阻遏物 2022/10/18
β-半乳糖苷酶 透过酶 转乙酰3酶2
2022/10/18
16
调节机理:
细胞中某一氨基酸或嘧啶的浓度发生改变
氨酰 – tRNA的浓度变化
核糖体在转录产物RNA上的结合位置不 同,使得RNA形成特定的二级结构 由RNA的二级结构判断基因能否继续转录
2022/10/18
17
3、降解物对基因活性的调节P252
葡萄糖效应或降解物抑制作用:细菌培养基中在 葡萄糖存在的情况下,即使加入乳糖、半乳糖等 诱导物,与其对应的操纵子也不会启动,这种现 象称为葡萄糖效应或降解物抑制作用。
这是通过阻止乳糖操纵子表达来完成的,这种 效应称为降解物抑制(catabolite repression)。
2022/10/18
35
(五)、cAMP与代谢物激活蛋白
葡萄糖
葡萄糖-6-磷酸
甘油 某些代谢产物抑制活性
腺苷酸环化酶
ATP
cAMP
编码
cAMP-CAP
Crp基因
代谢物激活蛋白 CAP
葡萄糖对其它糖的代谢抑制,是通过对 cAMP的抑制完成的。
2022/10/18
22
一、酶的诱导 ——
lac 体系受调控的证据
两种含硫的乳糖类似物:
异丙基巯基半乳糖苷
(IPTG)
巯甲基半乳糖苷(TMG)
E. coli 在不含乳糖的培养基生 长时,β-半乳糖苷酶含量极低;
分子生物学课件第七章 蛋白质的生物合成-翻译

2020/10/28
38
2020/10/28
39
原核生物翻译过程中核蛋白体结构模式:
P位:肽酰位 (peptidyl site)
A位:氨基酰位 (aminoacyl site)
E位:排出位 (exit site)
2020/10/28
40
蛋白质生物合成体系
mRNA、tRNA、rRNA n 氨基酸
大多数简并性表现在密码子的第三个核苷酸上,即 第一、二个核苷酸确定后,第三个核苷酸可变。
色氨酸
意义: 简并密码子越多,生物遗传的稳定性越大,
氨基酸出现频率越高
2020/10/28
18
(3)摆动性(wobble)
转运氨基酸的tRNA的反密码需要通过碱基互补与 mRNA上的遗传密码反向配对结合,但反密码与密码间 不严格遵守常见的碱基配对规律,称为摆动配对。
氨基酰-tRNA + AMP +PPi
2020/10/28
31
(2)氨基酰tRNA合成酶(amino acyl-tRNA synthetase,AARS)
存在于胞液中,催化一个特定的aa结合到相应的 tRNA分子上。
每种氨基酰tRNA合成酶对相应氨基酸以及携带氨 基酸的数种tRNA具有高度特异性,保证tRNA能够 携带正确的氨基酸对号入座。
UUU,UUG,UGU,GUU, GGG,GGU,GUG,UGG。 • U和G随机加入到三联体中,这样按比例各个位置 上进入U和G的概率不同,如氨基酸测定结果:
2020/10/28
7
• 如UUU:UGG=(555):(511)
= 25 : 1
• 同理UUU:UUG =5 :1,
• 根据检测结果推测:
分子生物学-13-4-第七章原核基因表达调控-Arb

因此只有在没有葡萄糖的时候,同时又有半乳糖的时候,启动子1才是开放的为什么gal 操纵子需要两个转录起始位点?(涉及半乳糖在细胞代谢中的双重功能)半乳糖两个作用: 可以作为唯一碳源供细胞生长; 与之相关的物质--尿苷二磷酸半乳糖(UDPgal )是大肠杆菌细胞壁合成的前体。
而启动子也有两个: galP1起始的转录——无内源葡萄糖、有外源半乳糖时进行,以保证碳源的供应。
galP2起始的转录——有内源葡萄糖、无外源半乳糖时进行,以保证细胞壁的合成需要。
生理功能(可以理解为生物学意义?)无论从必要性和经济性考虑,都要有一个不依赖于cAMP-CAP 的启动子(s2) 进行本底水平的组成型合成,以及一个依赖于cAMP-CAP 的启动子(s1),进行高水平的调节,这样既可以满足细胞最基本的需要(细胞壁),又可以满足在没有葡萄糖而有半乳糖时,细胞能够利用半乳糖进行生长。
进一步解释:gal P2是不依赖于cAMP-CRP 的,相反: cAMP-CRP 对gal P2还起到一种抑制作用,这是因为其与结合位点的结合,会影响到RNA 聚合酶对gal P2的利用。
因此教材上(page257)认为:只有S2活性完全被抑制时,(S1)的调控作用才是有效的。
7.4.2 阿拉伯糖操纵子(arabinose operon)araB 基因、araA 基因和araD, 形成一个基因簇,简写为araBAD三个基因的表达受到ara 操纵子中araC 基因产物AraC 蛋白的调控。
C 蛋白有三个结合位点O2、O1和 I 。
I BADCRPO2O1C结构基因调节基因P BADaraC 基因是araBAD 的调节基因L核酮糖激酶L阿拉伯糖异构酶L核酮糖-5-磷酸-4-差相异构酶结合到ara I 的时候,由于araBAD的启动子本身与ara I有部分重叠,另外还可以引起上游序列回折弯曲,使得AraC同时与O2结合,从而使CRP 聚合酶也不能结合到启动子上,araBAD基因不转录。
分子生物学技术-第七章 DNA的重组与转座

指不含任何结构基因的转座子,它是一种自主 的单位,仅带有介导自身移动所需要蛋白质的基 因,是最简单的转座子。
IS 一般由一段DNA序列(一个转座酶基因,不 带其它标记基因)和两端反向重复序列组成,重 复序列一般在15-25bp。
不同IS插入位点不同,但序列大多为5-9bp, 转位发生后插入位点成为IS两端的同向重复序列。
一、霍利迪结构(Holliday Structure)
霍利迪结构:它是DNA重组过程中的一个中间体, 重组时两个DNA双链体以四股DNA在连结点形成的十字 形DNA分子构象。
异源双链 (Hetero duplex):指在重组部位,每条双 链中均有一段DNA链来自另一个双链中的相对链。
分支迁移(Branch migration):一旦两个重组的 DNA分子形成霍利迪结构,交叉连接点很容易像拉链一 样移动,从而扩大异源双链体区域,这一过程叫分支迁 移。
2、复合式转座子
它是一类较复杂的转座子,由两个重复序列 夹着一个或多个结构基因组成。除了含有转位相 关的功能外,还带有一些其他基因,如抗药性基 因、抗重金属离子基因等。
它们都具有末端反向重复序列、可编码基因 和插入位点的正向重复序列
TnA转座子:
结构较复杂,长度约5000bp。两端没 有IS结构,仅含有两个末端反向重复序列, 一般约38bp。内部通常含有转位相关的基 因和抗药性基因等,插入位点约5bp。
第七章 DNA的重组与转座
重组分为4类:
同源重组:重组的DNA之间具有同源性 位点特异性重组:重组DNA之间不需要同源,但
是需要识别特异的DNA序列 转座重组:需要转座子
异常重组:不正常细胞加工的结果。
第七章-分子荧光法

第七章分子荧光分析法第一节概述物质的分子吸收一定的能量后,其电子从基态跃迁到激发态,如果在返回基态的过程中伴随有光辐射,这种现象称为分子发光(molecular luminescence),以此建立起来的分析方法,称为分子发光分析法。
物质因吸收光能激发而发光,称为光致发光(根据发光机理和过程的不同又可分为荧光和燐光);因吸收电能激发而发光,称为电致发光;因吸收化学反应或生物体释放的能量激发而发光,称为化学发光或生物发光。
根据分子受激发光的类型、机理和性质的不同,分子发光分析法通常分为荧光分析法,燐光分析法和化学发光分析法。
荧光分析法历史悠久。
早在16世纪西班牙内科医生和植物学家N.Monardes,就发现含有一种称为“Lignum Nephriticum”的木头切片的水溶液中,呈现出极为可爱的天蓝色,但未能解释这种荧光现象。
直到1852年Stokes在考察奎宁和叶绿素的荧光时,用分光计观察到它们能发射比入射光波长稍长的光,才判明这种现象是这些物质在吸收光能后重新发射的不同波长的光,从而导入了荧光是光发射的概念,并根据荧石发荧光的性质提出“荧光”这一术语,他还论述了Stokes 位移定律和荧光猝灭现象。
到19世纪末,人们已经知道了包括荧光素、曙红、多环芳烃等600多种荧光化合物。
近十几年来,由于激光、微处理机和电子学新成就等科学科术的引入,大大推动了荧光分析理论的进步,促进了诸如同步荧光测定、导数荧光测定、时间分辨荧光测定、相分辨荧光测定、荧光偏振测定、荧光免疫测定、低温荧光测定、固体表面荧光测定、荧光反应速率法、三维荧光光谱技术和荧光光纤化学传感器等荧光分析方面的发展,加速了各种新型荧光分析仪器的问世,进一步提高了分析方法的灵敏度、准确度和选择性,解决了生产和科研中的不少难题。
目前,分子发光分析法在生物化学,分子生物学,免疫学,环境科学以及农牧产品分析,卫生检验、工农业生产和科学研究等领域得到了广泛的应用。
分子生物学基础第七章真核基因表达的调控第三节真核基因表达转录水平的调控

第七章 真核基因表达的调控
第三节 真核基因表达转录水平的调控
一、真核基因转录与染色质结构变化的关系 DNA绝大部分都在细胞核内与组蛋白等结合成染色质, 染色质的结构影响转录,至少有以下现象: 1.染色质结构影响基因转录 在真核细胞中以核小体为基本单位的染色质是真核基 因组DNA的主要存在方式。DNA盘绕组蛋白核心形成核小体, 妨碍了与转录因子及RNA聚合酶的靠近和结合,使基因的 活性受到抑制。 2.组蛋白的作用 组蛋白H1及核心组蛋白共同参与核小体的组装与凝聚。 在特殊氨基酸残基上的乙酰化、甲基化或磷酸化等修饰, 可改变蛋白质分子表面的电荷,影响核小体的结构,从而 调节基因的活性。
第三节 真核基因表达转录水平的调控
图7-6 碱性螺旋-环-螺旋结构图
第三节 真核基因表达转录水平的调控
螺旋-转角-螺旋结构域是最早发现于原核生物中的一个关键因子, 该结构域长约20个aa,主要是两个α-螺旋区和将其隔开的β转角。 其中的一个被称为识别螺旋区,因为它常常带有数个直接与DNA序列 相识别的氨基酸。其结构如图7-3所示。
图7-3 螺旋-转角-螺旋结构及其与 DNA的结合
第三节 真核基因表达转录水平的调控
2.增强子 增强子是指能使基因转录频率明显增加的DNA序列。增强子的作 用有以下特点。 ①增强效应十分明显。一般能使基因转录频率增加10~200倍,有 的可以增加上千倍, ②增强效应与其位置和取向无关。 ③大多为重复序列。 ④增强效应有严密的组织和细胞特异性。说明只有特定的蛋白质 (转录因子)参与才能发挥其功能。 ⑤没有基因专一性,可以在不同的基因组合上表现增强效应。 ⑥许多增强子还受外部信号的调控,如金属硫蛋白的基因启动区 上游所带的增强子,就可以对环境中的锌、镉浓度做出反应。 ⑦增强子要有启动子才能控
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
b) 碱基异构式引起DNA复制的错配
(来源:分子生物学(2007),郑用琏,第321页)
碱基异构式引起DNA复制的错配
(来源:分子生物学(2007),郑用琏,第321页)
碱基异构式引起DNA复制的错配
正确配对 A(a) T(k)
G(k) C(a)
错误配对 A(a) C(i)
G(k) T(e)
A(i) C(a) G(e) T(k)
卫星搭载育种 太空蔬菜
(来源:不详)
微重力 高辐射 强射线
b) 非电离辐射—Ultra Violet light (U.V) ∧
---pyrimidine dimer (TT dimer )is generated by covalent links between adjacent TT
共价键
7.1. Point mutation 的类型与 机理 7.2.诱发突变 7.3. 保证遗传稳定的机制
7.4. 基因重组交换的分子机制
● conversion(取代)
transition(转换)
Py Py Pu
Pu
transvertion(颠换) Py Pu
● dNt deletion or insertion = 3x dNt ±n xAmino acid = 3x dNt Framshift
U.V.
…C T T A…
(来源:分子生物学(2007),郑用琏,第309页)
U.V.
deamination—oxidation
C
U A(a)
U.V. H2O H+ + OH-
C(a)
C(i) A(a)
U.V.
A(a, anti) G(k, syn)
C(a, anti) G(k, saynnti))
7.1.2.2.增变基因(mutatorgene )
but be wronged
w.t.
维持遗传的稳定性
mut.
随机引起其他各类基因的突变
增变基因类别;
修 DNApolymerase 相关基因
复
过
3’ 5’editing function mutation
Gene Mutation &
Crossing-Over
第7 章 基因突变与交换
7.2. 诱发突变
(Source:Lyer,V,The transcriptional program in the response of human fibroblasts to serum.Science 283(1 Jan 1999)f.1,p.83)
• 无效突变 Null mutation: 完全消除了基因功能的突变(缺失)
• 功能丧失型突变(loss-of-function mutation) 无效突变或其他阻止基因功能的突变
• 功能获得型突变(gain-of-function mutation) 突变使蛋白质获得新的功能
• 沉默突变(silent mutation) 没有明显表型效应改变的突变
7.2.1. 物理诱变
a) 电离辐射诱变;
Co60 (χ)( γ) ray) ray
(χ)( γ) ray穿透性
(外照射处理)
(α) (β) ray非穿透性
(内标记处理)
卫星搭载诱变; 高真空,强辐射,微重力
dNt电荷及结构改变
● 突变的表达类型 ---获得突变型是遗传学研究的重要前提 ---非条件型突变;allelein DNA level (RFLP, RAPD…) allelein phenotype (红花/白花,糯/非糯…) 条件型突变; 突变的表现= 突变基因型+ 诱导条件
(光,温敏感不育,Ts,su--…)
● 突变的表达类型
A(i, anti) A(a, syn) A(i, anti) G(k, syn)
G(e,i, anti) G(k, syn) G(e,i, anti) A(a, syn)
碱基异构式引起DNA的错配突变
A(a)
A(a)
C(i)
G(k)
C(ai)) T(k)
A(a, anti) T(k, anti)
第四,生物通过调节自身变异能力而适应环境的能力, 突变在进化中的作用比人们原先想象的要大得多相当巨。
● conversioneffect ---Samesense mut. ---Missense mut. ---Nonsense mut.
GAA(E) → GAG(E) GAA(E) → AAA(K) GAA(E) → TAA(stop)
南京大学田大成等发现的遗传突变新机制
Nature,doi:10.1038/nature07175,Dacheng Tian,Jian-Qun Chen
第一,基因组各区域的突变率很不相同,自发突变的数 量是由Indel的数量和密度所决定
第二,生物多样性最初变异来源主要是由Indel诱导产生 第三,自然选择在很大程度上是通过对Indel的选择而实 现
第7 章 基因突变与交换
Source:StanleyN. Cohen/Photo Researchers,Inc
Gene Mutation &
Crossing-Over
(Source:Lyer,V,The transcriptional program in the response of human fibroblasts to serum.Science 283(1 Jan 1999)f.1,p.83)
7.1.2. 突 变 发 生 的 机 理 (自发突变,诱发突变)
7.1.2.1. 自发突变
7.1.2.1.1. 碱基异构式引起DNA复制过程的错 误
a) 碱基异构式
A(amino) G(keto) T(keto)
A(imino) C(a)
C(i)
G(enol)
G(k)
G(e,i)
T(enol-2’) orT(enol-4’)
程
是 基
错配修复系统的基因
因
突
MCE (mismatch correctionenzyme)
变
的
重 DNA损伤修复系统基因
要
来 源
错配修复功能丧失
突变率升高
7.1.2.3 不对称交换 内源转座子 (Retro-transposon,Helitron)
Indel
Source:StanleyN. Cohen/Photo Researchers,Inc