哺乳动物毛色遗传
专题二遗传规律经典例题

B.F1高茎抗病植株的基因型有四种
C.F1中抗病植株与感病植株的比为3:1
D.F1抗病植株间相互随机传粉,后代抗病植株占8/9
4.某哺乳动物的毛色由常染色体上的一组等位基因B1、B2和B3控制,基因型与表现型如表所示。
基因型
B1B1
B1B2
B1B3
B2B2
B2B3
(4)家蚕中D、d基因位于Z染色体上,d是隐性致死基因(导致相应基因型的受精卵不能发育,但Zd的配子有活性)。能否选择出相应基因型的雌雄蚕杂交,使后代只有雄性?,请根据亲代和子代基因型说明理由:。
9.Ⅰ.下图甲表示某雄性动物的细胞分裂图像,其中①~④表示细胞中的染色体;图乙是某同学绘制的相关坐标曲线图。请回答下列问题:
(4)现已查明,B、b与D、d基因均位于10号染色体上,且不发生交叉互换。将一基因型为AABbDd的玉米植株进行测交,如果所结子粒的性状分离比为白色∶红色∶紫色=1∶1∶0,请在下方框中画出基因在染色体上可能的位置关系(只要画出与上述基因相关的染色体,用竖线表示染色体,黑点表示基因的位点,并标上相应的基因)。如果它们的基因在染色体上的位置关系不是如你所画,则测交后植株所结子粒的性状及分离比应为。
(1)图甲所示细胞的名称为,对应图乙的段。
(2)根据图甲推断,此动物正常体细胞最多有条染色体。
(3)理论上讲,图甲所示细胞分裂结束产生的4个精子,其染色体组成共有种。
(4)若A、a、B、b分别位于①②③④染色体上,而此细胞分裂结束最终产生了AB、aB、Ab、ab 4种精子,其原因之一是减数分裂中同源染色体的之间发生了交叉互换。
6.(8分)某农科院培育出新品种香豌豆(自花传粉,闭花受粉),其花的颜色有红、白两种,茎的性状由两对独立遗传的核基因控制,但不清楚花色性状的核基因控制情况,回答以下问题:
羊驼毛色相关基因研究进展的综述_邸文达 (1)

羊驼因其优质的毛纤维及其色彩多样的天然毛色越来 越受到重视。直接利用天然色,可免去深加工中漂白染色的 程序,节约成本,亦可减少对环境的污染。 羊驼毛色形成过 程复杂,但主要受控于遗传因素。基因主要通过调控黑色素 细胞的生成和分布来影响被毛颜色。掌握毛色遗传规律,发 掘控制毛色相关基因,科学育种,将有利于提高毛产量。 1 Agouti 基因在不同毛色羊驼皮肤组织中的表达
第8期
邸文达等:羊驼毛色相关基因研究进展的综述
85
驼 KIT 基因 intron18 核苷酸序列有 95%相同,因此可推测, 在 同 一 品 种 内 ,KIT 基 因 对 毛 色 影 响 的 原 因 不 一 定 是 由 于 intron18 缺失 4 个碱基导致 KIT 基因表达失调而致[9]。 5 AIF 基因在不同毛色羊驼皮肤组织中的表达
畜牧与饲料科学
Animal Husbandry and Feed Science
2012 ,33 (8 ):84-85
羊驼毛色相关基因研究进展的综述
邸文达,杨 川 (山西农业大学动物科技学院,山西 太谷 030801)
摘要:羊驼毛纤维颜色丰富,具有 22 种天然色,与其他哺乳动物一样,主要由遗传基因决定。 掌握控制羊驼毛色基因的
β-catenin 基因在黑色素细胞发育过程中, 参与神经嵴 的分化和成黑色素细胞的迁移, 并且在黑色素的转运过程 中也发挥作用。 Jouneau 等人已经证明,在黑色素细胞形成 过程中有 β-catenin 的表达 。 [11] 在于秀菊等研究 β-catenin 在不同毛色羊驼皮肤中的基因和蛋白表达时, 通过对 βcatenin 在羊驼皮肤组织的表达进行了 定 位 ,β-catenin 在表 皮、皮脂腺、根鞘部和毛乳头均有表达,在不同毛色羊驼皮 肤中的蛋白表达量不同, 在棕色羊驼皮肤组织中的蛋白表 达量远高于羊驼的表达量 。 [12] 7 Wnt3α 基因在不同毛色羊驼皮肤组织中的表达
广东省部分中学2023高中生物必修二第一章遗传因子的发现考点题型与解题方法

广东省部分中学2023高中生物必修二第一章遗传因子的发现考点题型与解题方法单选题1、若某哺乳动物毛色由位于常染色体上3对独立遗传的等位基因决定。
其中,A基因编码的酶A可使黄色素转化为褐色素,B基因编码的酶B可使褐色素转化为黑色素,D基因的表达产物能完全抑制A基因的表达;相应的隐性基因a、b、d的表达产物没有上述功能。
若用两个纯合黄色品种的动物作亲本进行杂交,F1均为黄色,F2中毛色表现型及比例为黄:褐:黑=52:3:9,则亲本的基因型组合是()A.AABBDDxaaBBdd或AabbDDxaabbddB.aaBBDDxaabbdd或AAbbDDxaaBBDDC.aabbDDxaabbdd或AabbDDxaabbddD.AAbbDDxaaBBdd或AABBDDxaabbdd答案:D分析:1 .分析题干可知,基因之间的关系为;A基因编码的酶可使黄色素转化为褐色素;B基因编码的酶可使该褐色素转化为黑色素:D基因的表达产物能完全抑制A基因的表达;故褐色对应的基因型为A_bbdd,黑色对应的基因型为A_B_dd,其他基因型均表现为黄色;2 .3对等位基因是位于常染色体上的、独立分配的基因,故三对等位基因满足基因的自由组合定律,F2中毛色表现型及其比例黄:褐:黑=52:3:9,褐色占3/64,褐色的基因型为A_bbdd,可推知3/64为3/4×1/4×1/4的结果,故F1的基因型为AaBbDd。
据分析可知,3对等位基因是位于常染色体上的、独立分配的基因,故三对等位基因满足基因的自由组合定律,F2中毛色表现型及其比例黄:褐:黑=52:3:9,不难发现,52+3+9=64=43,是(3:1)3的变形,可推导F1的基因型为AaBbDd,则亲本的基因型组合是AAbbDDxaaBBdd或AABBDDxaabbdd,ABC错误,D正确。
故选D。
2、某牵牛花植株与另一红花宽叶牵牛花(AaBb)杂交,其子代表型之比为3红花宽叶:3红花窄叶:1白花宽叶:1白花窄叶,此牵牛花植株的基因型和表型是()A.Aabb 红花窄叶B.AAbb 红花窄叶C.AaBb 红花宽叶D.aaBb 白花宽叶答案:A分析:基因分离定律的实质是:位于同源染色体的等位基因随着同源染色体的分开和分离。
猪的毛色遗传

猪 的毛色 遗传 是猪 的遗传 特性 之一, 一般 认为 毛色 与经济 性状 关系 不大, 不具 有直接 经济 价值 , 上研 究手 段和 方法 的限制 , 而很 长一 段时 间研 究较 加 因 少, 仅有 少量研究 , 且其 研究对象 仅分 布于数 量有 限的欧 洲和北美 品种 , 未发 现 研 究对 象有 亚洲 品种 。在二 十 世纪 5 0年代 前, 对猪 的毛 色遗 传 有 定 的研 一 究, p l m n 9 6 【首 次描述 了猪 的毛色遗 传, 后 的研 究主 要是在 1 4  ̄ S i l a (1 0 ) i j 此 95 1 5 年 发表 的 8篇系 列文 章 中 】 94 : 。近 年来 , 一些研 究 发现 毛色 与 猪的 生长速 度 、瘦 肉率及 对 疫病 的抵抗 力 等生 产 性 能也 有 直接 关 系。 。研 究毛 色 遗传 有利 于保持 品种 内各个 体 问的毛 色统 一 : 另外 , 色变 异还 可鉴 定某些 特 定的 毛 杂交育 种方案 成分, 在进行杂 交育 种时, 若能弄清 各种 毛色 的遗传规 律, 有利 于 获得育 种 目标所 确定 的或市场 所要 求的毛色 类型 : 同时又 发现毛 色是 种 可 利 用 的遗传 标记 , 在确 定杂 交 组合 、 品种纯 度 和 亲缘 关系 以及 评 价产 品质量 等 方 面均 有 一 定 用 途 。 1猪毛 色形 成 的生物 学 基础 猪 的毛色 是 由于黑 色素 的种类 和分 布情 况不 同而 造成 。在 哺乳动 物 的被 毛 中, 色素 有两 种,~ 种 为 以黑色 和 褐色 两种 形 式存 在 的 “ 黑 色素 ” 另 黑 . 真 : 种 是 以 黄色 和 红 色 两种 形 式 存在 的 “ 黑 色素 ” 褐 。黑 色素 是 一 种 结合 蛋 白质, 由酪氨 酸 酶 以及 与 之密 切相 关 的化学 物质 氧化 后 形成 的, 推测 黄色 和 红色可 能 是形 成黑色 素 过程 的提 前终 止而 产 生 。另外 , 素颗 粒 的浓 度和 分 色 布对 着 色 的变 化 也起 了很 大 的作 用 。 2猪 的毛 色类 型 猪 的毛色 类型很 多, 前面 已述, 主要 是 由于黑 色素 的种类 和分 布情 况不 同
哺乳动物毛色遗传

• 研究表明,微量元素在各种色素合成、代谢的调控网络中 扮演着非常重要的作用( Waterland et al.,2003) 。
• 铜是多酚氧化酶的组成部分,能促进酪氨酸转变成黑色素 ,对黑色素和棕色素形成具催化作用;锰对机体吸收铜有 一定的影响,这可在一定程度上影响动物的毛色形成( 角 根琴等, 2008)。
2.毛色形成的基础
黑色素细胞主要位于表皮和毛囊中黑色素包括真 黑色素( eumelanin ) 和褐黑色素( pheomelanin) 。 褐黑色素为溶于碱的圆形红色颗粒,能使皮肤或 毛发表现为黄色和红色; 真黑色素比褐色素难溶, 能使皮肤或毛发表现为褐色和黑色( 庞有志, 2008) 。
真黑色素和褐黑色素两种色素分布与比例的不同 造就哺乳动物不同的毛色( Cone et al.,1996; Hartmeyer et al.,1997) 。
– Ay 黄色,纯合时致死
– Aw 背部刺鼠毛,腹部黄白色 – A 刺鼠毛 – at 背部毛为褐色或黑色,腹部毛为淡黄色 – a纯合时为非刺鼠毛,呈全褐色或全黑色
B基因
• B基因:褐色基因。 • 有2个主要的等位基因B和b, B为黑色, b为棕色
C基因
• C基因:白化型基因,控制不同性质的色素氧化酶 的形成,共有5个复等位基因
2.毛色形成的基础
• 哺乳动物毛色是由体内色素决定的。虽然哺乳动 物体内的色素有很多种,但现有研究表明: 决定 哺乳动物毛色的物质基础主要是酪氨酸源性色素 ,即黑色素及其衍生物; 同时,这些色素的空间 分布和时空表达也影响到了哺乳动物毛色的深浅 和花纹组成( 里德等,1989; 张俊珍等,2006; 王 乐等, 2009a; 苗永旺等, 2009) 。
猪的杂交利用

猪的杂交利用一、质量性状的遗传和选择(一)、毛色遗传规律猪的毛色与其经济性状关系不大,但是品种的重要标志,在生产实践中,常根据猪的毛色变化来粗略鉴别其纯度。
猪的毛色之间有显隐性关系,其遗传符合孟德尔遗传规律。
因此,掌握毛色类型及其遗传规律具有重要意义。
猪的主要毛色类型有:1、白色:这类猪的被毛为全白色。
其品种很多,如长白猪、大白猪、哈白猪、上海白猪等。
白毛色猪与其他毛色猪交配,后代均为白色,因此,白色对其他毛色为显性,对野猪毛色也为显性。
2、纯黑色:被毛全为黑色。
这类猪种也很多,如我国的民猪、北京黑猪、内江猪、英国的巴克夏猪和美国的波中猪等。
黑色对棕红色为显性遗传,但因显性强度不同,分为完全显性和不完全显性。
完全显性遗传,如具有白环带的汉普夏猪黑毛色对棕红色的杜洛克是完全显性,我国多数的黑猪对棕红色猪也呈显性遗传;不完全显性遗传,如将有六白特征的巴克夏猪和波中猪与杜洛克猪杂交,其后代的毛色有红色和黑斑点,即所谓的虎斑毛色。
3、白环带:这类猪的腰部或颈肩部为白色,躯体两端为黑色,如我国的金华猪、宁乡猪以及美国的汉普夏猪等。
白环带毛色的猪与棕红色和黑色的猪交配,白环带趋向显性遗传,但白环带对我国某些黑猪为不完全显性。
4、花斑:其全身分布有大小不同的黑白花斑,如广东大花猪、北京花猪及比利时的皮特兰猪等。
5、棕红色其全身都为棕红色或棕黄色,如我国云南的大河猪、美国的杜洛克猪等,其遗传为隐性遗传,有的为不完全显性遗传。
(二)、耳型遗传规律猪的耳型和大小呈中间型遗传,垂耳对大型立耳呈不完全显性,用纯合立耳与纯合的垂耳猪杂交,其后代表现为半垂耳。
(三)、有害基因控制的遗传规律遗传疾病是指由基因突变或染色体畸变引起的某种形态缺陷、生理机能失常或生化紊乱的现象。
据统计,仔猪中患遗传缺陷的占1%。
1、猪应激综合征:肌肉丰满的猪在应激因子的作用下突然死亡,屠宰后肌肉为PSE 肉。
猪应激综合征呈常染色体隐性遗传,在某些育肥性状和胴体性状上,这种杂合体的性能优于显性纯合体的猪,在选种时常被选留下来,从而增加了氟烷基因频率。
遗传实验设计----某哺乳动物的毛色由位于常染色体上的三对等位基因决定

遗传实验设计某哺乳动物的毛色由位于常染色体上的三对等位基因决定,这三对等位基因控制毛色合成的关系如下图所示。
请回答下列问题:(1)某研究小组为研究相关基因是否遵循基因的自由组合定律,选择纯合的雌雄黄色个体作为亲本进行杂交实验,得F1后,再让F1中的雌雄个体随机交配,观察统计F2的表现型及比例。
①杂交亲本的基因型组合是。
(代表基因的字母按英文字母的先后顺序排列)。
②理论上,若F2中个体的表现型之比为黄色:褐色:黑色=,则说明这三对基因自由组合。
(2)另一研究小组通过基因敲除技术培育出不含D基因的该动物纯种个体,并利用它们进行上述杂交实验,经过多次重复实验,发现F2中个体表现型及比例接近黄色:褐色:黑色=4:1:7.①请针对出现该分离比的原因,提出一种合理的假说:。
②利用F2中的个体设计实验验证上述假说,写出实验步骤并推测实验结果。
实验步骤:。
实验结果:。
【解析】F2中个体的表现型之比为黄色:褐色:黑色=53:3:9。
(53+3+9=64,说明相关3对等位基因之间遵循基因的自由组合定律)应该是F1 AaBb产生的AB、Ab、aB、ab的四种配子中某种配子致死,4份黄色不变,说明致死的配子不可能是aB、ab。
若AB♀和♂配子都致死,则后代黑色占2/9,不符合题意;若AB♀或♂配子致死,则后代黑色占5/12,不符合题意;若Ab♀或♂配子致死,则F2中个体表现型及比例黄色:褐色:黑色=4:1:7,符合题意;【答案】(1)①AABBDD×aabbdd或AAbbDD×aaBBdd②53:3:9(2)①基因型为Ab的雌(或雄)配子致死②选择F2中褐色雌(或雄)个体与黄色雄(或雌)个体进行杂交,统计并观察后代的表现型子代个体的毛色全为黄色。
毛发颜色的形成 哈士奇基因遗传的秘密

毛发颜色的形成哈士奇基因遗传的秘密不同的品种的哈士奇,毛发的颜色和组成是不一样的。
形成哈士奇毛发颜色不同的基本原因就是哈士奇的遗传基因在起作用。
基因不同,所以哈士奇幼犬的毛发也不是相同的。
而且有的时候因为基因遗传的缘故,在买狗网身上所体现出来的方式也不一样。
像有一些颜色可以在哈士奇下一代中体现出来,而还有一些颜色需要在下两代或者好几代中才能显现。
这就是决定哈士奇毛发颜色的秘密--基因决定着哈士奇毛发的颜色。
举例说明纯色基因的遗传,比如一条携带两个黑色基因的纯毛哈士奇公犬与一条携带黑色和红色两个基因的哈士奇母犬交配的情况。
哈士奇公犬和哈士奇母犬均显示黑白毛色,因为哈士奇母犬的红色隐性基因被她的显性黑色基因掩盖了。
四个后代都遗传了两个纯色的基因,一个来自父亲一个来自母亲,但是来自父亲或母亲的哪一个基因则是自由选择的。
基因遗传的比例显示如下。
每条小狗都从父亲那遗传黑色基因,因为它们的父亲只有黑色基因。
它们从母亲那遗传要么黑色基因要么红色基因,都是百分之五十的机率。
所有的后代都显示黑白毛色,因为黑色显性基因掩盖了红色的隐性基因。
不可能分辨出哪个后代携带了红色隐性基因,因此都象它们的父母一样显示的是黑白毛色。
如果一条狗携带两个黑色基因与另一条携带两个红色基因的狗交配,它们的后代也将显示黑白色,因为它们都从父亲那遗传了黑色基因(父亲只有两个黑色基因可以提供)。
然后,所有的哈士奇价格后代都将携带一个隐性基因(红色基因)遗传自母亲(母亲只有红色基因可以提供)。
从表面上看不出这些后代是否携带红色基因,但如果知道它们的母亲是红色的,则他们一定携带一个隐性红色基因。
如果一条狗携带了一个黑色和一个红色的基因,与另一条同样携带一个黑色和一个红色基因的狗交配。
后代的毛色显示的比例如下图。
一条狗将显示红色,因为它携带两个红色隐性基因,一个来自它的父亲,一个来自它的母亲。
其它的后代将显示黑白颜色,尽管会有两条狗各自携带一个红色隐性基因。
猫的毛色遗传

【科普2】猫的毛色遗传——(转载,自己学习的,和大家共享)看到一些关于猫的配色的配色表。
但只有纯色的,无法找到所有颜色的配色表,请教过许多人都说没见过全花色的。
当时奇怪,我见到的猫猫分明什么颜色花纹,千姿百态什么都有。
如果不是纯色和纯色的配会出什么颜色呢,还有例如黑色和黑色会出蓝色,那为什么呢?出蓝色的机率又是多少了。
... 我都突然很想了解。
出于极大的好奇心,我想做一个所有花色相配都齐全的表。
由于无法找到现成资料,我就看了有关遗传学方面知识,查找关于猫的颜色基因,按照遗传的公式计算。
然后突然明白原来纯色表原来就是万能表。
所有我们眼中看到的千百花色的猫全部都遵守纯色表的规律。
了解了猫的毛色控制基因,所有花色的猫相配都能算出后代的颜色。
而且不是很深的学问,任何人都能自己计算得出来。
... 曾几何时一直以为猫的颜色不一定遗传自父母,可以遗传到祖父母或是更远,以为猫有颜色有无数种,了解之后才明白猫的颜色一定遗传自父母,控制颜色深浅和变化的隐性基因的才有可能遗传前几代。
大概就是说黄猫和黄猫不可能生出黑猫,只能生黄猫,也生不出红猫。
红猫和红猫可以生出黄猫,但却不可能生出不同色系的黑猫。
... 由于我不是猫的专业繁殖者,也不是遗传学专家。
关于猫的配色遗传资料是通过我自己整理,自己计算,所以误差有所难免。
这里的所有资料除非写明转贴的。
全部是纯个人的看法,不能说对,更不能说权威,所以请参考此网页的朋友只能作为参考意见,有不同看法欢迎指教,请大家共同探讨。
猫的颜色... 猫的颜色只有两种,就是红和黑。
除此之外没有别的颜色。
红色的淡化色是乳色。
黑色的淡化色是蓝色。
... 猫的颜色遗传是在性染色体上的。
... 高中生物课老师教过控制人的性别的系一对染色体。
猫同样一样。
公用 XY 表示,母用 XX 表示。
后代是从父母身上各拿一个字母。
如果在爸爸身上取了 Y ,在妈妈身上取了 X (从母亲处取只能是 X ),合起来就是 XY ,后代为男性。
3-毛色的遗传

鸡 的 羽 色 遗 传
芦花羽
即横斑羽, 即横斑羽,就是在有色 底上有白色带。 底上有白色带。由性连锁 基因B控制, 基因B控制,呈不完全显 性,有加性作用。公鸡为 有加性作用。 BB, BB,故其白带宽度比母鸡 大,因此公鸡的羽色比母 鸡淡。 鸡淡。B还使胫及喙部色 泽变淡, 泽变淡,因此公鸡的胫及 喙部比母鸡淡 纯种芦花鸡可自别雌雄 纯种芦花鸡可自别雌雄
叶黄素
畜禽机体内不合成,直接 从饲料中摄取得来,摄取 后储存在皮肤及体脂里
毛 色 的 遗 传
黑色素存在于黑素细胞 胞质中的黑素体中。黑 素细胞在胚胎发育过程 中由神经胚转移到身体 的其他部位,从而在这 些部位出现色素沉着。 黑素体在被毛生长过程 中通过细胞的胞吐作用 转移到被毛中,从而出 现色素的沉着。黑色素 贮积在皮肤的真皮层, 毛纤维的皮质层
毛色对产肉率产蛋率等无直接影响毛色是品种的主要特征之一是品种外貌的重要标志是最引人注目的品种特征对毛皮用家畜极为重要部分毛色还与致死基因或其他有害基因有关毛色的遗传不同畜种不同品种的毛色变化多端是因为畜禽的毛发和皮肤里含有色素的种类数量颗粒的形状及分布方式不同导致荣昌猪临高猪色素黑色素叶黄素真黑色素褐黑色素形成黑色或棕色为圆形红色颗粒形成黑色棕色褐色或黄色畜禽机体内不合成直接从饲料中摄取得来摄取后储存在皮肤及体脂里黑色素存在于黑素细胞素细胞在胚胎发育过程中由神经胚转移到身体的其他部位从而在这些部位出现色素沉着
婆罗门牛
猪 的 毛 色 遗 传
猪的毛色控制基因主要有鼠灰 )、扩散 )、显性白 (A)、扩散(E)、显性白(I) )、扩散( )、显性白( ) 和条带( )等多个系列。 和条带(B)等多个系列。
猪的毛色主要分为白色、黑色、褐色、红色、 猪的毛色主要分为白色、黑色、褐色、红色、花斑等
绵羊、犬、马、兔的毛色遗传

4座位D,基因d具有稀释毛色作用,使黑色变为青色而D则无此作用。 5座位E,该座位有4个复等位基因,E表现为黑色,E(m)为铁灰色,E(br)
为虎斑缟纹,e具有抑制黑色素作用。它们基因的显隐性关系为E > E(m) > E (br) > e.
6座位S,该座位有4个复等位基因,S为有色毛;S(i)出现爱尔兰或小白斑,
① 被毛,主要红色的有:枣骝、红栗(赤马)、红兔褐、红斑马、红青马等, 当母马被毛含有红毛时.其后代出红色被毛的机率就高.而且虽母马毛色 名称有别.但后代差异不大.也说明被毛遗传率要大于长毛。 ② 对于黑枣骝、黑克粟、绣黑马、深铁青马.被毛中部含有黑毛者.虽毛 色叫法上有区别.但其后代毛色上却互为衍生,没有明显的区别,进而说 明毛色遗传仍被毛为主。 所以在马匹育种固定毛色时.应以被毛颜色为基础.不应以人们毛分类 的叫法为依据。
的栗色。
(5)P座位,白斑基因P作用于背部出现白蹄, 显性基因则无此作用。 设特兰矮马
(7)其他座位,如si 、T、Bio、O等。
兔的毛色遗传
兔的毛色遗传与家兔育种,兔的毛色是一种重要的遗传标记,长期以来
人们对兔毛色遗传的研究非常重视,通过多种杂交试验,揭示了决定家兔毛色遗传 的一些基因座位及其遗传规律,并在家兔的育种和生产中得到广泛应用。
S(p)为不规则白斑;S(w)使白斑面积扩大至全身,它们基因的显性等级为S> S(i)> S(p)> S(w)。
7座位R,该座位等位基因的关系为不完全显性,基因型Rr为沙毛,显性或隐
性纯合时(RR 、rr)表现为其他毛色。
8座位M,显性基因M可使犬的白色被毛上出现斑纹,而隐性基因m无此作用。 9座位G,显性基因G存在时,可是出生的黑毛变为灰毛,而隐性基因g无此作
不同品系小鼠的毛色是如何形成的呢?
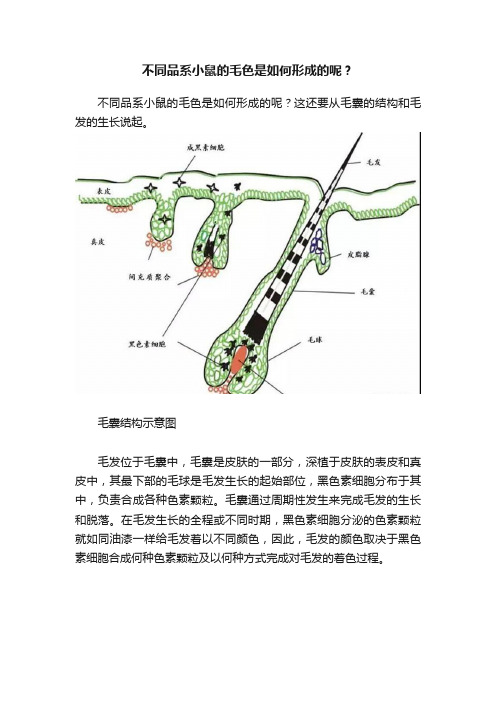
不同品系小鼠的毛色是如何形成的呢?不同品系小鼠的毛色是如何形成的呢?这还要从毛囊的结构和毛发的生长说起。
毛囊结构示意图毛发位于毛囊中,毛囊是皮肤的一部分,深植于皮肤的表皮和真皮中,其最下部的毛球是毛发生长的起始部位,黑色素细胞分布于其中,负责合成各种色素颗粒。
毛囊通过周期性发生来完成毛发的生长和脱落。
在毛发生长的全程或不同时期,黑色素细胞分泌的色素颗粒就如同油漆一样给毛发着以不同颜色,因此,毛发的颜色取决于黑色素细胞合成何种色素颗粒及以何种方式完成对毛发的着色过程。
真黑色素和褐黑色素的生物合成黑色素细胞合成的色素颗粒分为两大类,由两条不同的生产线来完成。
一条合成真黑色素(Eumelanin),其色素颗粒的颜色从黑色到棕色分布;另一条合成褐黑色素(Pheomelanin),其色素颗粒颜色从黄色到红色分布。
两条生产线的起始合成底物都是酪氨酸(Tyrosine),在酪氨酸激酶(TYR)的作用下转变成为多巴醌(DQ)---黑色素合成的中间体。
Balb/c 小鼠,图片来自Jax实验室网站如果酪氨酸激酶突变并丧失了生物活性,则色素颗粒无法合成,就形成白化小鼠。
常以大写字母C或Tyr来表示有活性的酪氨酸激酶,小写字母c或Tyrc表示无活性的酪氨酸激酶。
C对c为显性,根据孟德尔遗传,所有白化小鼠如Balb/c,FVB和ICR等该位点的两个等位基因均为无活性的酪氨酸激酶,简写为c/c。
C57BL(BLACK)、 C57BR(BROWN)在酪氨酸激酶有活性的前提下,真黑色素生产线上的色素颗粒成分又受酪氨酸相关蛋白(Tyrp1)基因调节。
Tyrp1有活性时,催化真黑色素生产线生产更多偏黑色的色素颗粒,携带该基因的小鼠毛发将被着以黑色,如C57BL小鼠(C57BL/6小鼠的祖系)。
如Tyrp1基因活性丢失,则真黑色素生产线将以合成棕色色素颗粒为主,携带该基因的小鼠毛色将表现为棕色,如C57BR小鼠。
常以大写字母B或Tyrp1代表该基因有活性,小写字母b或Tyrp1b代表该基因活性丢失,B对b为显性。
猪毛色遗传机制的研究进展

猪毛色遗传机制的研究进展张 建1,2,陈 伟1,王 慧1,曾勇庆*,1(1. 山东农业大学动物科技学院,山东 泰安 271018;2. 西藏林芝工布江达县农牧局,西藏 林芝 860200)近年来的一些研究发现,家畜的毛色与生长速度、瘦肉率及抗病力等重要经济性状有关[1]。
猪的毛色是重要品种特征之一,在养猪生产中具有多种作用,例如,毛色可以作为一种遗传标记,应用于评判品种的纯度、遗传稳定性、亲缘关系及在配套系培育中确定亲本的杂交组合类型[2]。
随着猪的杂种优势的广泛应用,为使商品猪具有理想的毛色,选择亲本品种的毛色也成为一个需要认真考虑的性状。
因而,揭示猪毛色的遗传机制及其影响毛色的相关基因的遗传规律,对于猪的育种具有重要意义[3]。
为了尽快提高猪的生产性能,许多国家都由过去封闭式的纯种选育改为采取开放式育种,导入某些优良基因,搞配套系生产,以最大限度地集中优良性状,但是,对毛色性状则未作严格的标准规定。
我国新引进的部分种猪,在纯繁中会出现少量毛色分离现象;其杂交后代,尤其与毛色不同的品种或品系的杂交后代会出现毛色分离,应属正常现象。
但是,作为育种场,在保持种猪高性能的同时,也应注意选择毛色一致性强的种猪,淘汰毛色分离严重的个体,提高品种的整齐度。
哺乳动物毛色的形成和遗传机制较复杂[4-8],是由一系列与黑色素细胞的种类与形成有关的生理、生化反应的最终结果。
猪的毛色虽然属于质量性状,由少数几对基因决定,但是,由于基因之间有上位效应等遗传互作效应,还有一些调控毛色的基因的最终作用尚未完全搞清楚,因而,猪的毛色的遗传十分复杂。
研究猪的毛色遗传的方法一般是采用杂交、自交和测交等方法,对后代群体(分离群体)进行毛色表型类型分组,并对各组个体数比例进行统计分析和遗传分析[10-12]。
1 猪的毛色类型及其形成机制1.1 猪的毛色类型猪的毛色有多种类型,可分为以下几种:野生型:特征是背部黑毛末梢有黄色横纹,且不同部位的颜色深度不同,野猪出生时有纵向条纹,以后会逐渐消失,如须野猪(bearded pig sus barbatus)、卷毛野猪(cebirfons)等。
绵羊犬马兔的毛色遗传专家讲座

基因,其完全显性突变基因 ,若为该基因杂合型时,则绵羊表现花斑, 若为纯合型时则表现体躯被毛白色而头部黑色。其对应等位基因无此 作用。
绵羊犬马兔的毛色遗传专家讲座
第5页
10斑点状阿卡拉曼位点L ,该位点有2个等位基因。其突变基因LAK
① 被毛,主要红色有:枣骝、红栗(赤马)、红兔褐、红斑马、红青马等, 当母马被毛含有红毛时.其后代出红色被毛机率就高.而且虽母马毛色名 称有别.但后代差异不大.也说明被毛遗传率要大于长毛。
② 对于黑枣骝、黑克粟、绣黑马、深铁青马.被毛中部含有黑毛者.虽毛 色叫法上有区分.但其后代毛色上却互为衍生,没有显著区分,进而说明 毛色遗传仍被毛为主。
绵羊犬马兔的毛色遗传专家讲座
第15页
绵羊犬马兔的毛色遗传专家讲座
第16页
1.2 家兔毛色基因座位及其基因关系
家兔毛色遗传十分复杂,现已证实控制家兔毛色基因最少有 8个基因座位(或系统),这些基因座位都位于常染色体上.当前 在性染色体(x和 Y)上还没有发觉控制毛色基因,也就是说毛色 遗传与性别无关。在决定毛色基因座中,多数基因座存在有复 等位基因系列,各基因座上等位基因及其基因间显隐性关系见 表 1。常见家兔品种毛色及其对应基因型见表 2 。
因B对栗色基因b显性。
(3)着色基因座位C,除马属动物外,几乎全部哺乳
动物,都存在粉红色眼和皮肤,这种症状叫白化症。它 是C座位隐性基因作用结果。
(4)不完全显性座位D。 (5)色素扩延座位E,有3个复等位基因,E(D)能使黑色
素扩散,E能促进黄色素扩散。E无此作用,它们显性等 级为E(D) >E >e。
萨米尔 公马,黑骝毛
基因遗传会造成哈士奇毛色变化

基因遗传会造成哈士奇毛色变化常听到有些人讨论,两只红色的哈士奇配出来,小狗是什么颜色的?黑的配红的,小狗又是什么颜色的?为什么有的有白围脖?为什么带天窗?为什么会有奶牛斑?这些问题的答案,都是有其规律可循的,简单的说;就是基因遗传。
在这里,不谈太深奥的东西,尽量能以较通俗易懂的语句,来阐述一些基本概念,例如天窗;白脖;奶牛等等通俗的用语。
在进入正题之前,一些关于遗传的基本原则和专有名词,还是要提一下,否则没法继续。
基因(Gene):孟德尔在遗传分析中所提出的遗传因子,如决定豌豆种子的圆形(R)和皱形的(r)等。
这些因子现代的术语来说就是基因。
基因位于染色体上,是具有特定的核苷酸(主要是DNA)片段,是储存遗传信息的功能单位。
等位基因(Allele):在染色体上占据相同位置的两个不同的基因基因座(Locus):基因在染色体所处的位置。
特定的基因在染色体上都有其特定的座位。
每个基因座上,有两个等位基因。
显性基因(DominantGene):在杂合状态中,能够表现出表型效应的基因,以大写字母表示。
隐性基因(RecessiveGene):在杂合状态中,不表现出表型效应的基因,以小写字母表示。
纯合体(Homozygore):基因座上有两个相同的基因。
例如BB,bb杂合体(Heterozygous):基因座上有两个不同的基因,例如Bb,Dd.犬有39对;78个染色体,交配时,精子和卵子各带来自父代和母代的39个染色体,随机又有序的结合成子代的39对染色体。
例如父代控制颜色的基因座上的两个等位基因为Bb,而母代控制颜色的基因座上的两个等位基因为BB,而所谓有序,是说父代控制颜色的基因,只会和母代控制颜色的基因结合,不会去和控制尾位的基因结合。
所谓随机是说精子所带的基因可能是B,也可能是b.随机的和卵子所带的B结合,子代基因的结果可能是BB或Bb有了这些基本概念,继续谈正题。
关于犬类毛色的基因形态有很多种,如下表:其中关于哈士奇的,比较重要的有三种基因形态:基因座B 的等位基因对;基因座D的等位基因对和基因座S的基因系列:以下分别说明:位于基因座B为两个控制颜色的不同的等位基因;B和b.B 为显性基因,其表型为黑色,b为隐性基因,其表型为棕色或俗称的红色。
猪毛色的遗传和选择

猪毛色形成的生物学基础
• 猪显性白毛色的形成
– 黑色素细胞前体物如能在生皮节及上皮细胞之间的通道正常迁移, 将可产生黑色素细胞 – 黑色素细胞前体物表达肥大/干细胞生长因子受体,此受体对应的配 体为肥大细胞生长因子,当肥大细胞生长因子缺乏时,黑色素细胞前 体物便不能迁移。
• KIT基因编码肥大/干细胞生长因子受体,因此,KIT基因发生变异,必 然扰乱肥大细胞生长因子在生皮节和上表皮通道间的扩散,黑色素细 胞将不能成活,最终导致毛根中缺乏黑色素细胞及其前体物,毛色表现 为白色。
猪毛色基因的研究进展
1. 白色(I)基因座
– I基因在白色品种(如大白猪、长白猪)中通常是纯合的,I基因抑制黑 色素、黄色素的形成。有色品种如巴克夏、波中猪、大黑猪、杜 洛克、皮特兰以及有色的中国品种都是隐性基因的纯合体(ii)。 – 在I基因座除了I和i等位基因外,还发现了Id、Ip、im等位基因,其显性 系列为I>Id>Ip>i>im。Id对I呈隐性,但当存在Ep时,Id与I同样对色素 的形成有抑制效应,如IdiEpEp基因型产生隐性暗白,当存在E基因 时,Id产生灰杂色(grey-roan), 实际是黑毛或白毛的混生,与牛、马的 杂色基因同源。 – 等位基因Ip是最近才提出来的,原因是当白色品种(大白、长白)和欧 洲野猪或中国黑猪杂交时,F1偶尔可观察到黑斑,Johansson等(1992) 认为这是由于I基因座有一个Ip等位基因的作用,这个等位基因在欧 洲白色品种分离频率很低,主要存在于斯堪的纳维亚国家的白猪。
猪毛色基因的研究进展
1. 白色(I)基因座
–现代品种中白色是最主要的毛色。早在1906年,Spillman 通过泰姆华斯与约克夏杂交试验,确认了白色对有色显 性。Wright和Lush(1923)提出白色受单一显性基因控制 的假说。Hetzer(1945)证实了这个假说,并把这个基因称 为I(抑制色素)基因, 现已证明I基因等同于KIT基因,且I 基因和毛色扩展基因E基因座是相互独立的,进一步证 实了上述假说。
哺乳动物毛色形成机制与影响因素

哺乳动物毛色形成机制与影响因素吴宇婷【摘要】哺乳动物拥有丰富多彩的毛色,对于多种多样的动物毛色,影响其形成的可能原因较多,遗传物质与环境的相互作用不可忽视.本文针对调控黑色素细胞形成及褐黑色素、真黑色素合成的相关基因,分析了哺乳动物毛色形成的可能机制,并对研究不同毛色动物的同种动物遗传差异的方法进行了讨论.【期刊名称】《四川动物》【年(卷),期】2011(030)006【总页数】5页(P1003-1007)【关键词】毛色;形成机制;褐黑色素;真黑色素【作者】吴宇婷【作者单位】四川大学生命科学学院,成都610064【正文语种】中文【中图分类】Q959.8;Q954哺乳动物斑斓的毛色组成了这个世界神奇而又丰富的色彩。
不同的毛色已成为一些动物亚种(或品种)区分的标志之一。
例如,猪的毛色可以大致分为纯黑、纯白、红棕、污白、野生型、花猪等多种毛色类型(方美英等,2002)。
牛、鼠、马等也有多种多样的突变毛色及固定毛色。
那么,究竟是什么原因导致了丰富多彩的毛色,又是什么决定着黑色与红棕色的转变呢?本文对多种哺乳动物毛色形成机制加以综述,就黑色素和真黑色素的合成以及调控黑色素细胞形成相关的基因、环境因素等对动物毛色形成原因进行初步分析,以期对哺乳动物毛色成因进行探讨。
哺乳动物毛色是由体内色素决定的。
虽然哺乳动物体内的色素有很多种,但现有研究表明:决定哺乳动物毛色的物质基础主要是酪氨酸源性色素,即黑色素及其衍生物;同时,这些色素的空间分布和时空表达也影响到了哺乳动物毛色的深浅和花纹组成(里德等,1989;张俊珍等,2006;王乐等,2009a;苗永旺等,2009)。
黑色素细胞主要位于表皮和毛囊中(Thierry et al.,2005)。
黑色素包括真黑色素(eumelanin)和褐黑色素(pheomelanin)。
褐黑色素为溶于碱的圆形红色颗粒,能使皮肤或毛发表现为黄色和红色;真黑色素比褐色素难溶,能使皮肤或毛发表现为褐色和黑色(庞有志,2008)。
探索基因奥秘:鸡羽色的遗传之旅

探索基因奥秘:鸡羽色的遗传之旅在自然界的缤纷画卷中,动物的羽色无疑是最为引人注目的一笔。
尤其是家禽中的鸡,其羽毛颜色的多样性一直是遗传学研究的热点。
近期,一项关于鸡羽色遗传机制的研究,为我们揭开了隐藏在色彩背后的遗传密码。
本篇文章将深入探讨这一领域的最新发现,并尝试从更广阔的视角审视鸡羽色遗传学的深远意义。
羽色遗传:从基因到表型的奥秘鸡的羽毛颜色是由多种基因共同作用的结果,这一点已在学术界达成共识。
然而,具体是哪些基因在幕后操控着这一复杂的遗传过程,一直是研究者们努力探索的问题。
近期,一项发表在《基因》杂志上的研究为我们提供了新的线索。
研究者们通过对比罗得岛红鸡(RIR)和白来航鸡(WL)的杂交后代,发现了控制羽色的特定基因变异。
特别是,一个位于SOX10基因上游的7.6 kb非编码区的缺失,被证实与浅色黄羽鸡(LY)的表现型直接相关。
基因与环境:羽色变异的双重奏羽色的形成不仅受到遗传因素的影响,环境因素同样扮演着重要角色。
家鸡的祖先——红原鸡,其羽色相对单一,而家鸡则展现出多样的羽色。
这种变化,部分可以归因于人工选择和环境适应。
在不同的饲养环境下,某些羽色可能更有利于鸡的生存和繁殖,从而在自然选择的过程中被保留下来。
此外,人工选择也在一定程度上塑造了鸡的羽色多样性,人们根据自己的喜好和需求,选择性地繁育具有特定羽色的鸡种。
基因组学:解锁羽色遗传的金钥匙随着基因组学技术的发展,我们对鸡羽色遗传机制的理解日益深入。
全基因组关联研究(GWAS)和高通量测序技术的应用,使得研究者能够在更精细的层面上探索羽色的遗传基础。
通过比较不同羽色鸡只的基因组数据,科学家们能够识别出与特定羽色表现型相关的遗传变异。
这些发现不仅丰富了我们对家鸡遗传多样性的认识,也为家禽的遗传改良提供了新的分子标记。
遗传多样性:生物进化的见证鸡羽色的多样性是生物进化过程中的一个缩影。
在漫长的进化旅程中,基因的变异和自然选择共同作用,塑造了今天我们所见的多样生命形态。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
• 研究表明,微量元素在各种色素合成、代谢的调控网络中 扮演着非常重要的作用( Waterland et al.,2003) 。
• 铜是多酚氧化酶的组成部分,能促进酪氨酸转变成黑色素 ,对黑色素和棕色素形成具催化作用;锰对机体吸收铜有 一定的影响,这可在一定程度上影响动物的毛色形成( 角 根琴等, 2008)。
2.毛色形成的基础
真黑色素与褐黑色素产生的调控模式示意图( Thierry et al.,2005; 张俊珍, 2006)
2.毛色形成的基础
• 黑色素存在于黑素细胞胞质中的黑素体中。 – 胚胎发育过程中,黑素细胞在由神经胚转移到 身体的其他部位,从而决定色素沉着的部位。 – 在被毛生长过程中,黑素体通过细胞的胞吐作
4.其他因素对毛色的影响
—其他环境因素对于毛色的影响
• 研究表明,氧化酪氨酸和苯丙氨酸合成真黑色素、褐黑色 素的酶系均对温度和性激素比较敏感( 张俊珍等2006) 。 • 体内的激素水平是受多方面综合因素影响的,包括光照、 气味、温度、海拔、年龄,甚至是否有异性在附近等。同一 种动物的毛色可能由于一些环境因素的不同而有差异,甚至 可以由于人类的饲喂方式不同而导致毛色的不同表达( 王乐 等, 2009b)。
用转移到被毛中,从而出现色素的沉着。
– 黑色素贮积在皮肤的真皮层,毛纤维的皮质层
3、影响毛色的主要基因
•对小鼠的研究表明,超过127 个基因座影响毛色的 形成过程,哺乳动物至少有A、B、C、D、E和S等主 要基因影响不同毛色的形成( Thierry et al.,2005; 张俊珍, 2006)。
白化松鼠
白化刺猬
白化猩猩
D基因
D基因:淡化 基因,包括D 栗色+DD 和d两个等位 基因,基因型 DD不淡化其 他毛色、Dd 栗色+Dd 有一定程度的 淡化,dd则很 大程度地淡化 其他毛色,见 栗色+dd 图示
栗 色
淡 褐 色 奶 酪 色
E基因
E基因:色素分化 德国牧羊犬中存在E和e两个等位基因 基因,决定整个 被毛黑色素扩散 或不扩散的基因。 这个位点上已知 的等位基因及显 性等级是:E m (黑嘴筒)— E (纯黑色)—e br (灰白色和棕色 条纹)—e(黄 色)。
S基因
S基因:斑点基因, 控制斑点的有或 斑点狗:黑白花斑 无。主要包括S和 s两个等位基因, 基因型S-无斑点、 ss出现斑点。除白 荷斯坦奶牛:黑 化外,S基因可与 白花斑 其他所有毛色同 时出现,如黑白 西门塔尔牛:黄白 斑、红白斑等, 花斑、 举例 淡红白花斑
4.其他因素对毛色的影响
—微量元素在不同毛色中含量不同对毛色形成的影 响
哺乳动物(宠物)的毛色遗传
主要内容
1. 动物毛色的意义
2. 毛色形成机制
3. 影响毛色的主要基因 4. 其他因素对毛色的影响 5. 宠物毛色的观赏价值
1.动物毛色的意义
毛色是品种的主要特征之一 躲避天敌、利于生存繁衍 是最引人注目的品种特征 对毛皮用家畜极为重要 观赏价值 方便动物生产管理(鸡自别雌雄) 部分毛色还与致死基因或其他基因有关
• 显性等级为 C> cch > cm > ch >c
– C为野生型,允许色素表达,一般为黑色 – cch青紫蓝
– cm黑貂毛色,基本色黑色,体侧及大腿部毛色很淡
– ch喜马拉雅型,为热敏感型,表型为全身大部分白毛, 只有在温度较低的体端如耳朵、鼻尖为黑毛 – c白化型albinos,阻止色素表达,cc缺乏色素,即白化
C基因
C基因:喜 马拉雅基 因型ch ch, 含此基因 型的动物 多作为宠 物出售 Himalayan mouse Siamese cat
Himalayan rabbits
长毛猫
喜马拉雅猫 体重:约3kg 身长:约18cm 原产于英国起源于1935年, 属人工繁殖的品种,与喜马 拉雅山无关。 喜马拉雅猫具有波斯猫与暹 (xian1)罗猫的血统,有可 爱的外表,具有双重性格。 有时温柔,有时比较活泼, 很恋家。 非喜马拉雅毛色
同时,由前面所述两种黑色素的调控模式可知,环境的细微 变化也会对黑色素的表达、毛色的变化造成一定程度的影响 。
5.宠物毛色的观赏价值
—以贵宾犬为例
贵宾的基本颜色主要是由两个“基因座”中,即“B基因 座”及“E基因座”。 BBEE/BBEe/BbEE/BbEe(毛色=黑色) bbEE/bbEe(毛色=咖啡色) BBee/Bbee/bbee(毛色=白色、奶油色、杏色或红色)
5.宠物毛色的观赏价值
最近北美非常流行的Parti-color poodle(乳牛 斑纹),因为不是单一色彩,所以还是不被认可 的颜色,但是因为非常美丽,所以在北美有很多 专业犬舍专门繁殖Parti-poodle。
Thanks!
的喜马拉雅猫
白化基因隐性纯合cc
• 白化牛 • 白化猪等
• 白化虎
• 白化猩猩 • 白化响尾蛇 • 白化企鹅 • 等等
• 在自然界中也有很多动物的毛色或羽色是白色的,如北极 狐、北极熊、大天鹅和白鹭等,它们并不是白化动物,其 白色的体色是由显性基因的正常表达。 • 而白化动物是一对隐性基因纯合子的产物,虹膜大多为红 色,往往还同时携带着其他对其自身不利的因素,如怕光 、眼球震颤、皮癌等,另外在自然界中也容易为天敌所发 现而受到攻击,所以比正常个体难于存活。不过,在人工 饲养的动物中,白化现象却很常见如白兔、小白鼠、大白 鼠、白马和白玉鸟等,它们是在长期人工精心选育和保护 下培育而成的,能够正常地繁衍后代。
• 白化动物大多发现在爬行类、鸟类和哺乳类动物中,其中 以哺乳类中发现的种类较多,人类也有这样的个体,属于 白化病,俗称为“天老儿”。 • 白化动物在我国古籍中和民间多有记载,如《史记•五帝 本纪》中关于白熊的记载、《魏略辑本》中关于白麋的记 载和《白蛇传》等民间传说,清朝宫廷画家、意大利人郎 世宁还依据宫中所藏的贡品,创作了很多有白化动物的画 。据说湖北神农架一带是白化动物出现最多的地区, 1987年曾发现白龟,全身为白色,只有双眼鲜红,颈部 透明,很像一个雕刻的艺會 死亡。 非常少见的白化蟒蛇
白化海豚
白 化 长 颈 鹿
白化梅花鹿
白化鹿
白化袋鼠相当罕见, 现在仅有几只都在澳 洲的动物园里,
由于白化病人的通病: 怕晒, 所以动物园护理员每 天都会在它们身上涂 抹防晒霜。
阿尔卑斯白化大角羊
白化狮子 白 化 鼬 鼠
2.毛色形成的基础
黑色素细胞主要位于表皮和毛囊中黑色素包括真 黑色素( eumelanin ) 和褐黑色素( pheomelanin) 。 褐黑色素为溶于碱的圆形红色颗粒,能使皮肤或 毛发表现为黄色和红色; 真黑色素比褐色素难溶, 能使皮肤或毛发表现为褐色和黑色( 庞有志, 2008) 。
真黑色素和褐黑色素两种色素分布与比例的不同 造就哺乳动物不同的毛色( Cone et al.,1996; Hartmeyer et al.,1997) 。
– Ay 黄色,纯合时致死
– Aw 背部刺鼠毛,腹部黄白色 – A 刺鼠毛 – at 背部毛为褐色或黑色,腹部毛为淡黄色 – a纯合时为非刺鼠毛,呈全褐色或全黑色
B基因
• B基因:褐色基因。 • 有2个主要的等位基因B和b, B为黑色, b为棕色
C基因
• C基因:白化型基因,控制不同性质的色素氧化酶 的形成,共有5个复等位基因
A基因
A基因:鼠灰色基因,也称刺鼠
毛基因。为毛色的野生型,即常见 的野兔毛型,其表型称agouti,表 现为动物背部的毛被每根毛有三个 色带,毛基部是污黑色、近端部是 黄褐色、顶端是黑色,见下图:
野兔
Black hair
Agouti hair
A基因
• A基因:共有5个复等位基因,显性等级为 Ay > Aw > A > at >a
2.毛色形成的基础
• 哺乳动物毛色是由体内色素决定的。虽然哺乳动 物体内的色素有很多种,但现有研究表明: 决定 哺乳动物毛色的物质基础主要是酪氨酸源性色素 ,即黑色素及其衍生物; 同时,这些色素的空间 分布和时空表达也影响到了哺乳动物毛色的深浅 和花纹组成( 里德等,1989; 张俊珍等,2006; 王 乐等, 2009a; 苗永旺等, 2009) 。