一、核糖体的形态结构和类型
第9章_核糖体

第9章核糖体第一节核糖体的类型和结构核糖体的模式图核糖体是合成蛋白质的细胞器,几乎存在于一切细胞内。
核糖体是一个颗粒状的结构,主要成分是蛋白质和RNA。
核糖体RNA成为rRNA,蛋白质称为r蛋白,蛋白质含量约占40%,RNA约占60%,r蛋白分子主要分布在核糖体的表面,而rRNA则位于内部,二者靠非共价键结合在一起。
电镜下,是无包膜的电子致密颗粒,略呈圆形或椭圆形,平均直径在150~250A。
核糖体由大、小两个亚单位组成。
大亚基略呈梨形,中心有一条中央管。
直径为230A,沉降系数为60S。
其上有与氨酰-tRNA 结合的位置,还含有转肽酶活性部位。
小亚基呈碟盘状,大小为230A×120A,沉降系数为40S,其上有蛋白质合成启动因子结合位点、起始氨酰-tRNA结合部位和mRNA结合位点。
电镜下,核糖体常成群呈丛状或螺旋状存在,与mRNA结合,构成多聚核糖体(polyribosome)。
附着于内质网上的称附着核糖体(bound ribosome),主要合成输送到细胞外的分泌性蛋白、膜嵌入糖蛋白、可溶性驻留蛋白和溶酶体蛋白等。
散在于胞质中的称游离核糖体(free ribosome),主要合成组成细胞本身所需的结构性蛋白质。
糖核体的大小两个不同的亚基,在不进行蛋白质合成时,它们是分开的,游离存在于细胞质中。
只是在进行蛋白质合成时才结合在一起。
原核生物和真核生物的核糖体成分的比较原核细胞的核糖体为70S,真核细胞线粒体和叶绿体内的核糖体也近似于70S,但除了这两个细胞器,真核细胞内的核糖体均为80S。
原核生物核糖体由约2/3的RNA及1/3的蛋白质组成。
真核生物核糖体中RNA占3/5,蛋白质占2/5。
真核细胞糖核体的沉降系数为80S。
大亚基为60S,小亚基为40S。
小亚基含有由一种18S的 rRNA 和33种蛋白质;大亚基含有5S、5.8S及 28S 三种rRNA 和约49种蛋白质。
tRNA结合部位1. tRNA的三叶草结构受体臂(acceptor arm)主要由链两端序列碱基配对形成的杆状结构和3′端末配对的3-4个碱基所组成,其3′端的最后3个碱基序列永远是CCA,最后一个碱基的3′或2′自由羟基(—OH)可以被氨酰化。
第11章 核糖体

15
E.coli核糖体小亚单位中rRNA与r蛋白的相互关系示意图 线条表示相互作用及作用力的强(粗线)与弱(细线) (引自Alberts et al,1989)
16
16SrRNA的一级结构是非常保守的 16SrRNA的二级结构具有更高的保守性:臂环结构(stemloop structure) rRNA臂环结构的三级结构模型
100s
120s
10
对核糖体结构进行精细分析的必
要手段是:获得高质量的核糖体晶
二、核糖体的结构
体,然后进行X射线衍射分析
1、对核糖体的X射线衍射图谱分析表明:
每个核糖体有供tRNA结合的3个位点(A、P、E),横跨大小亚
单位的结合面;
11
1、对核糖体的X射线衍射图谱分析表明:
每个核糖体有供tRNA结合的3个位点(A、P、E),横跨大小亚
化的rRNA进行核糖体的重组装,显示核糖体中r蛋白与 rRNA的结构关系。
₪同一生物中不同种类的r蛋白的一级结构均不相同 ₪不同生物同一种类r蛋白之间具有很高的同源性,并在进化上
非常保守。
₪蛋白质结合到rRNA上具有先后层次性。 ₪核糖体的重组装是自我装配过程
13
70S核糖体的小亚单位中rRNA与全部的r蛋白关系的空间模型 对rRNA,特别是对16SrRNA结构的研究
第十一章 核糖体
第一节 核糖体的类型与结构
第二节
多聚核糖体与蛋白质的合成
1
1953年Robinsin和Brown用电镜在植物细胞中发现; 1955年Palade在动物细胞中观察到;
核糖体是合成蛋白质的细胞器,其唯一的功能是按照mRNA的指令
由氨基酸高效且精确地合成多肽链。
9 第九章 核糖体

第九章 核糖体1第 节 第一节 核糖体的类型与结构核糖体的基本类型与成分 核糖体的结构 核糖体蛋白质与rRNA的功能分析2发现史: Robinson&Brown(1953)发现于植物细胞, Palacle(1955)发现于动物细胞 Roberts(1958)建议命名为核糖核蛋白体(ribosome), 简称核糖体。
简称核糖体 核糖体功能:是合成蛋白质的细胞器,其唯一的功能是按照 mRNA的指令由氨基酸高效且精确地合成多肽链。
mRNA的指令由氨基酸高效且精确地合成多肽链 形态:颗粒状,无被膜,25 nm 化学成分:rRNA(60%)和r蛋白(40%) 存在:真核,原核,线粒体,叶绿体 存在:真核 原核 线粒体 叶绿体 类别:附着核糖体,游离核糖体 数量:不定3Free and membrane-bound ribosomes4一、核糖体的基本类型与成分两种基本类型的核糖体:5核糖体聚合与解聚 聚 解聚大,小亚基 单核糖体 二聚体1 mMMg2+浓度10 mM核糖体大小亚基常游离于细胞基质 中,当小亚基结合mRNA后,大亚 基才与其结合,形成完整的核糖体。
6二、核糖体的结构7研究方法核糖体的重组装 - 离子交换树脂可分离纯化各种r蛋白 - 纯化的r蛋白与纯化的rRNA进行重组装,显示其结构关系 双功能的交联剂和双向电泳分离可用于研究r蛋白在结构上的相互关系 电镜负染色与免疫标记技术结合,研究r蛋白在核糖体的亚单位上的定位816S rRNA结构 16S rRNA的一级结构是非常保守的; 16S rRNA的二级结构具有更高的保 守性: 臂环结构 (stem-loop structure) rRNA臂环结构的三级结构模型。
3’ Minor 5’ Central 3 3’ Major9核糖体蛋白质与rRNA的功能核糖体上有一系列与蛋白质合 成有关的结合位点与催化位 点 ①mRNA的结合位点 ②新掺入的氨酰-tRNA的结合 位点——氨酰基位点(A位点) ③延伸中的肽酰-tRNA的结合 位点——肽酰基位点(P位点) ④肽酰转移后即将释放的 tRNA的结合位点 ——E位点(exit site)10③ ④②①如何判断核糖体中的A、P是两个分开的独立位点?的独立位点核糖体蛋白质与rRNA的功能⑤肽酰tRNA从A位点转移到肽酰P位点相关转移酶(即延伸因子EF-G)的结合位点⑥肽酰转移酶的催化位点肽酰转移酶的催化位点r蛋白or rRNA?实验证据:①无法确定哪一种蛋白具有催化功能,在E. coli中核糖体蛋白突变甚至缺失对①无法确定哪一种蛋白具有催化功能在E coli蛋白质合成并没有表现出“全”或“无”的影响②多数抗蛋白质合成抑制剂的突变株,并非由于r蛋白的基因突变而往往是②多数抗蛋白质合成抑制剂的突变株并非由于rRNA基因突变③在整个进化过程中rRNA的结构比核糖体蛋白的结构具有更高的保守性肽酰转移酶的催化位点Noller用蛋白酶和SDS处理E.Coli的50S亚单位,23S rRNA rRNA的主要功能:①具有肽酰转移酶的活性②为tRNA提供结合位点(A位点、P位点和E位点)③为多种蛋白质合成因子提供结合位点④在蛋白质合成起始时参与同mRNA选择性地结合以及在肽链的延伸中与mRNA结合p g⑤核糖体大小亚单位的结合、校正阅读(proofreading)、无意义链或框架漂移的校正以及抗菌素的作用等都与rRNA有关r蛋白质的主要功能关于r蛋白功能有多种推测,主要有:有多种推测主要有①对rRNA 折叠成有功能的三维结构是十分重要的②在蛋白质合成中某些r蛋白可能对核糖体的构象起“微调”作用③在核糖体的结合位点上甚至可能在催化作用中, r蛋白与rRNA共同行使功能第二节多聚核糖体与蛋白质的合成定义:多个核糖体串连在一条mRNA分子上定义多个核糖体串连在一条二、蛋白质的合成的基本过程链的起始⏹⏹链的延伸⏹链的终止消耗2ATP 第一步反应第二步反应3’-CCA-OHE = 氨酰-tRNA合成酶SD序列(SD sequence)(q)IRES进入位点(一)肽链的起始基与①30S小亚基与mRNA的结合起始因子(initiation factors,IF):IF1, IF2, IF3②第一个氨酰-tRNA进入核糖体第个氨酰大亚基小亚基组合③大基小基合(二)肽链延伸①氨酰位点tRNA进入A②形成肽键③转位④脱氨酰-tRNA的释放tRNA(三)肽链的终止核糖体沿着☐mRNA移动,如果进入A位的是终止密码子(UAA、UAG、UGA),终止合成☐终止需要释放因子(release factor, RF),与tRNA非常相似,能进入A位。
核糖体(ribosome)

多聚核糖体的解聚:是指多聚核糖体分散为单体,失去正
常有规律排列,孤立地分散在胞质中或附在粗面内质网膜
细
上。一般认为,游离多聚核糖体的解聚将伴随着内源性蛋
胞 生
白质生成的减少。脱粒是指粗面内质网上的核糖体脱落下 来,分布稀疏,散在胞质中,RER上解聚和脱离将伴随外 输入蛋白合成。
物
正常情况下,蛋白质合成旺盛时,细胞质中充满多聚核糖
核 糖 体
头部 基部 小亚基
基部
头部
平台
mRNA
a
中央突
谷
柄
嵴
大亚基
中央突 嵴
5
核糖体大小亚基的结合和解离依细胞的生理状 态和Mg²+浓度变化
细胞的生理状态
当细胞在合成蛋白质时,大小亚基结合
细
当细胞合成蛋白质合成结束时,大,小亚基解离
胞 生
Mg²+浓度
物
当Mg²+浓度大于0.001mol/L时,大,小亚基结合成
体
另外,一些药物,致癌物可直接抑制蛋白质合成的不同阶
段,有些抗苔素,如链霉素、氯霉素、红霉素等对原核与
真核生物的敏感性不同,能直接抑制细菌核糖体上蛋白质
的合成作用。有的抑制在起始阶段,有的抑制肽链延长和
终止阶段,有的阻止小亚基与mRNA的起始结合,四环素
抑制氨基酰-tRNA的结合和终止因子,氯霉素抑制转肽酶,
一个中间停靠点,只是作暂时的停
留。当E位点被占据之后,A位点
同氨酰tRNA的亲和力降低,防止
了氨酰tRNA的结合,直到核糖体
准备就绪,E位点腾空,才会接受下
一个氨酰tRNA。
a
11
小亚基的功能
细
将mRNA结合到核糖体上,并稳定mRNA与核糖体的结合;
第十一章 核糖体

第十一章核糖体核糖体是一种核糖核蛋白颗粒(ribonucleoprotein particle),是细胞内合成蛋白质、没由膜包被的细胞器,其功能是按照mRNA的信息将氨基酸高效精确地合成蛋白质多肽链。
因为富含核苷酸,1958年Roberts建议把这种颗粒命名为核糖蛋白体,简称核糖体(ribosome)第一节核糖体的类型与结构一、核糖体的基本类型与化学组成:生物界有两种基本类型的核糖体:一种是原核细胞核糖体;另一种是真核细胞核糖体。
两种核糖体都有两个大小不同的亚基(subunit)组成,每个亚基都含有rRNA和蛋白质。
原核细胞核糖体沉降系数为70S,相对分子质量为2.5*106,易解离为50S与30S的大小亚基。
真核细胞核糖体沉降系数为80S,相对分子质量为4.8*106,易解离为60S与40S的大小亚基。
rRNA中的某些核苷酸残基被甲基化修饰,甲基化常发生在rRNA序列较为保守的区域。
核糖体大小亚基常常游离于细胞基质中,只有当小亚基与mRNA结合后打牙祭才与小亚基结合形成完整的核糖体。
肽链合成终止后,大小亚基解离,又游离于细胞质基质中。
二、核糖体的结构结构与功能的分析方法表明:(1)离子交换树脂可分离纯化各种r蛋白。
(2)核糖体中r蛋白与rRNA的结构关系:纯化的r蛋白与纯化的rRNA进行核糖体的重组装的过程中,某些蛋白质必须首先结合到rRNA上,其他蛋白才能装配上去,即表现出现后层次。
(3)双功能的交联剂和双向电泳分离:可用于研究r蛋白在结构上的相互关系。
(4)电镜负染色与免疫标记技术结合:研究r蛋白在核糖体的亚单位上的定位。
(5)对rRNA,特别是对16S rRNA结构的研究已十分成熟:①16SrRNA的一级结构是非常保守的②16SrRNA的二级结构具有更高的保守性③16SrRNA可以分为四个结构域:中心结构域,5'端结构域,3'端结构域和主结构域。
蛋白质合成过程中很多重要步骤与50S核糖体大亚单位相关:(1)依赖延伸因子Tu(EF-Tu)的氨酰tRNA的结合;(2)延伸因子G(EF-G)介导的转位作用;(3)依赖于起始因子2的fMet-tRNA的结合;(4)依赖于释放因子的蛋白合成终止作用;(5)应急因子与核糖体结合产生阻断蛋白合成等。
细胞生物学(第五版)-第10章-核糖体精选全文完整版

二、蛋白质的合成
又称蛋白质的翻译,是细胞中最复杂、最精确的生 命活动之一。蛋白质合成需要各种携带氨基酸的 tRNA、核糖体、mRNA、多种蛋白质因子、阳离子 及GTP等的参与
蛋白质合成分为三步:
起始(Initiation)包括核糖体与mRNA 结合,形 成起始复合物,其中含有第一个氨酰-tRNA。
仅发现在哺乳动物成熟的红细胞 等极个别高度分化的细胞内没有 核糖体,线粒体和叶绿体中也含 有核糖体。 核糖体是细胞最基本的不可缺少 的结构。
核糖体是一种不规则颗粒状的结构,其主 要成分是RNA和蛋白质,直径约25 nm 核糖体蛋白分子主要分布在核糖体表面, 核糖体RNA(rRNA)位于内部,二者靠共价 键结合在一起。
甲基转移酶催化形成的。
30S小亚基与mRNA的结合需要 起始因子(initiation factor,IF)的 帮助。 这些起始因子仅位于30 S亚基上。 一旦30 S亚基与50 S亚基结合形 成70S核糖体后便释放。 起始因子的主要作用:帮助形成 起始复合物。 原核细胞有3种起始因子: IF1、IF2和IF3。
主要包括4个步骤: 1、氨酰-tRNA进入核糖体A位点的选择 2、肽键的形成 3、转位(translocation) 4、脱氨酰-tRNA的释放。
1.氨酰-tRNA在核糖体A位点的入位
起始的tRNAiMet占据P位点, 核糖体接受第2个氨酰-tRNA进 入A位点,这就是肽链延伸的 第一步。 为了有效地结合A位点,第二 个氨酰-tRNA必须与有GTP的 延伸因子(elongation factor, EF)EF-Tu结合形成复合物氨酰 -tRNA·EF-Tu·GTP。
三、核糖体蛋白质与rRNA的功能
核糖体上具有一系列与蛋白质合成有关的结合位 点与催化位点
核糖体

结合,使其不再缠结而便于作模板——去螺旋稳定蛋白
(HDP)。
与 复 制 有 关 的 另 外 两 种 酶
拓扑异构酶
拓扑异构酶I :切断DNA双链中的 一股,使DNA解链旋转 时不致缠结,待张力解 除后又把切口封闭。 拓扑异构酶II :稳定螺旋结构;当 复制完毕时,使着丝 粒处连锁着的两个 DNA分子分离。
:保证真核细胞内线 端粒酶(端粒末端转移酶) 性DNA的复制进行得 彻底和完善。
真核细胞DNA复制特点:
1. 碱基互补配对 2. 半保留复制 3. 复制的方向性 4. 复制是不连续的 复制子(replicon) , 复制叉(replication fork) 先行链和后随链 冈崎片段 5. 多个复制子双向复制 6.复制的不同步性 7.复制的引物:RNA 片段
DNA复制过程显示复制的不连续性、先行链和后随链
5’ 3’
O O P OOHO
3’ 5’
DNA连接酶
ATP
ADP
5’ 3’
O O P OO-
3’
5’
DNA复制 DNA连接酶
* rRNA的结构
* rRNA的功能 参与组成核蛋白 体,作为蛋白质生物 合成的场所。
* rRNA的种类(根据沉降系数)
真核生物
5S rRNA 28S rRNA 5.8S rRNA 18S rRNA
原核生物
5S rRNA 23S rRNA 16S rRNA
逆转录
(二)、遗传信息的翻译
携带某种遗传信息的mRNA转录出来后 经加工剪接,从细胞核进入细胞质,再与核 糖体大、小亚基以及甲硫氨酸tRNA结合 形成起始复合体, 蛋白质合成开始
第十一章 核糖体

• 根据核糖体存在的方式不同
任何核糖体都是由大小两个亚基组成
30S/40S
50S/60S
70S/80S
核糖体大小亚基的结合
核糖体的存在与Mg2+有关
•
[Mg2+]<1mM
• 70S
50S+30S
二聚化 [Mg2+] ≥ 1mM
• 100S
•
[Mg2+]<1mM
• 80S
60S+40S
二聚化 [Mg2+] ≥ 1mM
很难确定哪一种蛋白具有催化功能: 在E.coli中核糖体蛋白突变甚至缺失对蛋白 质合成并没有表现出“全”或“无”的影 响。
第一节 核糖体的类型与结构
rRNA的活性 多数抗蛋白质合成抑制剂的突变株,
并非由于r蛋白的基因突变而往往是 rRNA基 因突变。
在整个进化过程中rRNA的结构比核糖 体蛋白的结构具有更高的保守性。 由此推断: rRNA在蛋白质合成中可能具有重 要作用。
择性地结合以及在肽链的延伸中与mRNA结 合;
核糖体大小亚单位的结合、校正阅读 (proofreading) 、 无 意 义 链 或 框 架 漂 移 的 校 正、以及抗菌素的作用等都与rRNA有关。
• 120S
• 细胞质中的核糖体常以游 离的亚基形式存在。
第一节 核糖体的类型与结构
一、核糖体的基本类型与成分 2、主要成分
r蛋白质:占1/3,核糖体表面 rRNA: 占2/3,核糖体内部
原核生物与真核生物核糖体成分的比较
• 成分特点: r蛋白多为单拷贝;
如:细菌(E.coli.)r蛋白中L4/L12有4拷 贝、S6有 2拷贝,其余的r蛋白均为单拷贝。
11-14核糖体

~49种蛋白质
A
18S RNA
~33种蛋白质
5S RNA
5S RNA
第三节 核糖体的生物发生与自组装
真核细胞核糖体rRNA几乎都是在染色体的核仁 组织区(nucleolus organizer region, NOR) 内转录合 成的。
18S rRNA+ 33r蛋白质
•rDNA
45S rRNA
(前体分子)
+Mg++ —Mg++
二聚体 120S
核糖体以游离的形式存在于细胞质中,以可附着在内质网上。
第二节 核糖体的基本类型与 化学组成
一、核糖体的基本类型 原核细胞核糖体
类 种 真核细胞质核糖体
叶绿体核糖体
真核细胞器核糖体
线粒体核糖体
二、核糖体的化学组成
核糖体的类型 原核细胞核糖体 真核细胞核糖体 真核细胞器核糖体
Termination
终止包括两步:
1、RF与mRNA终止信号结合,激活转肽酶; 2、水解肽链和 tRNA间的键,新合成肽链、tRNA、
mRNA离开核糖体,后者即解离成大、小亚基, 进入下一轮反应。
(三)合成的蛋白质类型
结构蛋白: 又称内源性蛋白(endogenous protein),主 要由游离核糖体合成,多分布细胞基质中, 是用于细胞本身或参与组成细胞自身结构 的蛋白质,是细胞内代谢所需的蛋白质。
•rRNA - ribosomal RNA —Is what the ribosomes are made of
•tRNA - transfer RNA —Brings the amino acids to the mRNA to string them together to make a protein
第一节:核糖体的类型与结构
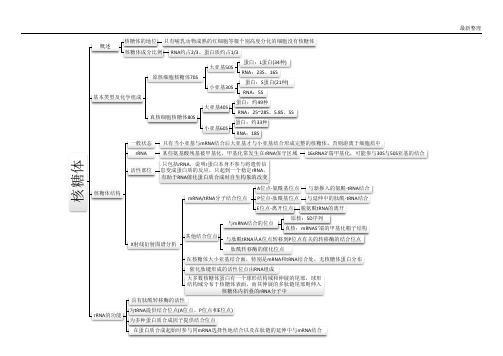
核糖体概述核糖体的地位只有哺乳动物成熟的红细胞等极个别高度分化的细胞没有核糖体核糖体成分比例RNA 约占2/3、蛋白质约占1/3基本类型及化学组成原核细胞核糖体70S大亚基50S蛋白:L 蛋白(34种)RNA :23S 、16S 小亚基30S蛋白:S 蛋白(21种)RNA :5S 真核细胞核糖体80S大亚基40S蛋白:约49种RNA :25~28S 、5.8S 、5S 小亚基60S蛋白:约33种RNA :18S核糖体结构一般状态只有当小亚基与mRNA 结合后大亚基才与小亚基结合形成完整的核糖体,否则游离于细胞质中rRNA某些氨基酸残基被甲基化,甲基化常发生在rRNA 保守区域16sRNA3'端甲基化,可能参与30S 与50S 亚基的结合活性部位只包括rRNA ,说明r 蛋白本身不参与将遗传信息变成蛋白质的反应,只起到一个稳定rRNA ,有助于RNA 催化蛋白质合成时自生构象的改变X 射线衍射图谱分析mRNA/tRNA 分子结合位点A 位点-氨酰基位点与新掺入的氨酰-tRNA 结合P 位点-肽酰基位点与延伸中的肽酰-tRNA 结合E 位点-离开位点脱氨酰tRNA 的离开其他结合位点与mRNA 结合的位点原核:SD 序列真核:mRNA5'端的甲基化帽子结构与肽酰tRNA 从A 位点转移到P 位点有关的转移酶的结合位点肽酰转移酶的催化位点在核糖体大小亚基结合面,特别是mRNA 和tRNA 结合处,无核糖体蛋白分布催化肽键形成的活性位点由RNA 组成大多数核糖体蛋白有一个球形结构域和伸展的尾部,球形结构域分布于核糖体表面,而其伸展的多肽链尾部则伸入核糖体内折叠的rRNA 分子中rRNA 的功能具有肽酰转移酶的活性为tRNA 提供结合位点(A 位点、P 位点和E 位点)为多种蛋白质合成因子提供结合位点在蛋白质合成起始时参与同mRNA 选择性地结合以及在肽链的延伸中与mRNA 结合多核糖体与蛋白质的合成多聚核糖体定义多个核糖体串联在一条mRNA 上形成的聚合体串联核糖体距离相邻核糖体中心距离30~35nm 相邻核糖体核苷酸距离:80个合成速度与mRNA 长度关系多肽合成速度提高的倍数与结合在mRNA 上的核糖体数目成正比mRNA 缩短,多聚核糖体的大小及核糖体的数目也成比例减少蛋白质合成原核与真核比较原核细胞中,mRNA 合成的同时,核糖体就结合到mRNA 上进行转录翻译过程(原核)肽链的起始30S 小亚基与mRNA 的结合识别mRNA 的起始密码子AUG-SD 序列位置:位于起始密码子AUG 上游5~10个碱基处,其他AUG 不具备作用:与核糖体小亚基16SrRNA3'端的碱基互补结合起始因子结合IF1与30S 亚基A 位点结合,协助30S 亚基与mRNA 的结合,并防止氨酰-tRNA 错误进入核糖体的A 位点IF2一种GTP 结合蛋白,协助第一个氨酰-tRNA 进入核糖体IF3能防止核糖体50S 亚基提前与小亚基结合,并有助于第一个氨酰-tRNA 进入核糖体,并调节核糖体动态平衡以及30S 亚基与mRNA 结合能力方面发挥重要作用。
第十二章 核糖体(ribosome)

L11-rRNA复合物的三维结构 (引自Porse et.al.,1999)
蛋白质的合成
又称蛋白质的翻译,是细胞中最复杂、 最精确的生命活动之一。 蛋白质合成需要各种携带氨基酸的tRNA、 核糖体、mRNA、多种蛋白质因子、阳离 子及GTP等的参与。
原核生物的蛋白质合成
(一)、 翻译起始
(1) 30S小亚基与mRNA 的结合:IF1、 IF2、 IF3、 (2)第一个氨酰-tRNA进入核糖体 携带有甲酰甲
第九章
核糖体(ribosome
核糖体的类型与结构
多聚核糖体与蛋白质的合成
2009年诺贝尔化学奖
三位科学家英国剑 桥大学文卡特拉 曼· 拉马克里希 南、美国科学家 托马斯· 施泰茨 和以色列科学家 阿达· 约纳特因 “对核糖体的结 构和功能的研究” 而获得2009的诺 贝尔化学奖。
贡献
都采用了X射线蛋白质晶体学的技术,标 识出了构成核糖体的成千上万个原子。 这些科学家们不仅让我们知晓了核糖体 的“外貌”,而且在原子层面上揭示了 核糖体功能的机理。
• ③医学和医药上的回收、分离和提纯。
• ④作为有机合成中的固体酸碱催化剂。 • ⑤食品及生物制品的脱色。
核糖体的重组装:将不同来源的核糖体大小 亚基、或不同亚基的 rRAN 或蛋白质重新组合, 形成杂合的核糖体后研究其功能的实验方法。 同一生物中不同种类的r蛋白的一级结构均不 相同,在免疫学上几乎没有同源性。 不同生物同一种类 r蛋白之间具有很高的同源 性,并在进化上非常保守。
免疫标记技术是指用荧光素、酶、放射性 同位素或电子致密物质等标记抗体或抗 原进行的抗原抗体反应。此技术优点很 多 : 特异、敏感、快速、能定性和定量 甚至定位,且易于观察。
电镜负染色
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
沉降系数(sedimentation coefficient,S) coefficient, 沉降系数( 颗粒在单位离心力场中粒子移动的速度。 颗粒在单位离心力场中粒子移动的速度。
为离心前粒子离旋转轴的距离; X1为离心前粒子离旋转轴的距离;X2为离 心后粒子离旋转轴的距离; 心后粒子离旋转轴的距离;ω是旋转角 速度
4.核糖体六个活性部位: 4.核糖体六个活性部位: 核糖体六个活性部位
结合部位: mRNA 结合部位:小亚 基上, mRNA结合 基上,与mRNA结合 A部位 :大亚基上,接 大亚基上, 受氨基酸―tRNA ―tRNA位 受氨基酸―tRNA位 部位: 小亚基上, P部位: 小亚基上,释 tRNA位 放tRNA位 肽基转移酶部位: 肽基转移酶部位:大亚 基上, 基上,催化肽键形成 GTP酶位 大亚基上, 酶位: GTP酶位:大亚基上, 移位A 移位A 到P 部位:大亚基上, E部位:大亚基上,新 生肽链出口位
一、核糖体的形态结构和类型
1.核糖体的形态大小 1.核糖体的形态大小 颗粒状,无膜包被( 颗粒状,无膜包被(非 膜性细胞器) 膜性细胞器)。 大小:15大小:15-25nm 游离于细胞质基质或 附着于内质网上。 附着于内质网上。 是细胞中合成蛋白质 的场所。 的场所
附着核糖体 核仁 游离核糖体
附着核糖体 示核糖体分布
(1)密码子 的方向性: 的方向性: 5′→3′ (2)密码子 的简并性与 兼职” “兼职” (3)密码子 的通用性 (4)密码子 是不传密码的特征 3.遗传密码的特征
二、核糖体与多肽链的合成
氨基酰氨基酰-tRNA 合成 酶具有高度的专一 性。 每一种氨基酰 -tRNA 合成酶只能 识别一种相应的 tRNA。 tRNA。 tRNA 分子能接受相 应的氨基酸, 应的氨基酸, 决定 于它特有的碱基顺 序, 而这种碱基顺 序能够被氨基酰序能够被氨基酰tRNA 合成酶所识别。
核糖体的形成
形成位置: 形成位置:核仁 形成过程: 形成过程:核仁 组织中心的rDNA 组织中心的rDNA rRNA基因 基因, 为rRNA基因, rDNA转录形成 rDNA转录形成 rRNA; rRNA; 蛋白质 由胞质经核孔进 入核仁区, 入核仁区,与 rRNA组装为大 组装为大、 rRNA组装为大、 小亚基, 小亚基,再经核 孔进入胞质。 孔进入胞质。
中心法则
DNA传递给 生物的遗传信息从 DNA传递给 复制 mRNA的过程称为转录。 mRNA的过程称为转录。 的过程称为转录 DN A 根据mRNA链上的遗传信 转录 根据mRNA链上的遗传信 mRNA 反转录 息合成蛋白质的过程, 息合成蛋白质的过程, R N A 蛋白质 翻译 被称为翻译和表达。 被称为翻译和表达。 复制 1958年 Crick将生物遗传信息的这种传递 1958 年 Crick 将生物遗传信息的这种传递 方式称为中心法则
(一)氨酰- tRNA合成(图示:酵母丙氨酰氨酰- tRNA合成 图示:酵母丙氨酰合成( tRNA 结构) 结构)
氨基酸 的活化 氨酰氨酰 tRNA的 tRNA的 合成
(二)肽链合成的起始(initiation) 肽链合成的起始(
小亚基附着 与mRNA上 上 甲酰甲硫氨 酰-tRNA识别 识别 AUG 起始复合物 的形成
三.原核细胞(Prokaryotic )和真核细胞 原核细胞( Eukaryotic)核糖体 (Eukaryotic)核糖体 化学组成比较
rRNA分子内部碱基配对形成许多短的双链区, rRNA分子内部碱基配对形成许多短的双链区, 分子内部碱基配对形成许多短的双链区 并形成螺旋状,非配对区形成环状或泡状; 并形成螺旋状,非配对区形成环状或泡状;共 同折叠成复杂的三维结构,组成核糖体骨架。 同折叠成复杂的三维结构,组成核糖体骨架。
二、核糖体的聚合和解离
Mg2 10mmol/L mmol/L时 1.当Mg2+为1~10mmol/L时,大、小亚基 聚合成单核糖体。 聚合成单核糖体。 Mg2 小于1mmol/L时 2.当Mg2+小于1mmol/L时,单核糖体解离 为大、小亚基。 为大、小亚基。 3.当Mg2+大于10mmol/L 10mmol/L时 3.当Mg2+大于10mmol/L时,两个单核糖 体结合成二聚体。 体结合成二聚体。
2.核糖体的类型:游离核糖体(合成细胞结构蛋白, 2.核糖体的类型:游离核糖体(合成细胞结构蛋白, 核糖体的类型 和附着核糖体(合成分泌蛋白、 分化低细胞内发达 )和附着核糖体(合成分泌蛋白、 膜受体、溶酶体蛋白,分泌功能旺盛, 膜受体、溶酶体蛋白,分泌功能旺盛,分化程度高的 细胞内发达 )。
RNA主要构成核糖体的骨架,将核糖体串联起来, RNA主要构成核糖体的骨架,将核糖体串联起来, 主要构成核糖体的骨架 并决定其定位
多聚核糖体的作用:一条mRNA上可有多个核 多聚核糖体的作用:一条mRNA上可有多个核 mRNA 糖体进行蛋白合成,提高了蛋白合成效率。 糖体进行蛋白合成,提高了蛋白合成效率。
3.核糖体的结构与组成 大亚基+ 3.核糖体的结构与组成:大亚基+小亚基 核糖体的结构与组成:
大小亚基一般以游离 状态存在, 状态存在,只有当小 亚基和mRNA mRNA结合后大 亚基和mRNA结合后大 小亚基才结合, 小亚基才结合,形成 完整的核糖体 核糖体是一种动态结 构,当参与翻译过程 大小亚基结合, 时,大小亚基结合, 蛋白质合成结束, 蛋白质合成结束,大 小亚基解体
思考题
核糖体的六个活性部位? 1. 核糖体的六个活性部位 ? 2. 真核细胞与原核细胞核 糖体的化学组成有什么不同? 糖体的化学组成有什么不同? 什么是遗传密码、 3. 什么是遗传密码 、 密码 遗传密码的特征? 子?遗传密码的特征? 4.氨基酸的活化过程
(四)肽链合成的终止及核糖体的释放 核糖体移行 至终止密码, 至终止密码 即终止密码 出现于A位 出现于 位, 肽链合成终 止; 大小 亚基解 离
蛋白质合成过程: 蛋白质合成过程:
原核细胞和真核细胞核糖体上蛋白质的合成
原核 细胞 转录 和翻 译的 地点、 地点、 时间 与真 核细 胞有 何不 同?
核糖体的形成示意图
第二节核糖体的功能: 第二节核糖体的功能:参与蛋白质的合成 一、遗传密码 1.遗传密码: 1.遗传密码: mRNA 遗传密码 分子中碱基排列顺序 2.密码子: mRNA分 2.密码子: mRNA分 密码子 子中三个相邻的碱基 决定一种氨基酸, 决定一种氨基酸,故 称其为三联体密码或 密码子。 密码子。
核糖体的功能
合成蛋白质的机器 游离核糖体:合成结构蛋白, 游离核糖体:合成结构蛋白,分化低细胞内发 达 ,如细胞内代谢酶、红细胞内血红蛋白、 如细胞内代谢酶、红细胞内血红蛋白、 组蛋白、核糖体蛋白、肌球蛋白等。 组蛋白、核糖体蛋白、肌球蛋白等。 附着核糖体:合成分泌蛋白,分泌功能旺盛, 附着核糖体:合成分泌蛋白,分泌功能旺盛, 分化程度高的细胞内发达,如肽类激素、抗体、 分化程度高的细胞内发达,如肽类激素、抗体、 消化酶类等, 消化酶类等, 溶酶体酶也是附着核糖体合成 的。
(三)肽链的延伸(Elongation): 肽链的延伸(
氨酰-tRNA进入 位 进入A位 氨酰 进入 肽键形成
肽链的延伸:核糖体继续沿5‘—3‘移动并添加氨基 肽链的延伸:核糖体继续沿5‘—3‘移动并添加氨基
酸到肽链上 移位: 移位:核糖体 沿mRNA5’-3’ 方向移动一个 密码子 A位上的肽基 位上的肽基 酰-tRNA移位 移位 到P位 位
几十种蛋白质(每种一份)通过与rRNA相互识别结 几十种蛋白质(每种一份)通过与rRNA rRNA相互识别结 合在rRNA骨架上,构成一个严格有序的超分子结构。 rRNA骨架上 合在rRNA骨架上,构成一个严格有序的超分子结构。
蛋白质
示蛋白质与rRNA的结合 的结合 示蛋白质与
50
32
原核生物与真核生物核糖体成分的比较