酶的活性部位
生物化学 核酸名词解释

1、Ribozyme:具有高效特异催化功能的RNA。
2、自杀性底物:Kcat型不可逆抑制剂不但具有与天然底物相似的结构,而且本身也是酶的底物,可被酶催化而发生类似底物的变化。
因此称之为“自杀性底物”3、酶的活性部位(活性中心):与底物接触并且发生反应的部位就称为酶的活性中心,也称为酶的活性部位。
4、变构酶又称别构酶,酶分子的非催化部位与某些化合物可逆地非共价结合后,引起酶的构象的改变,进而改变酶的活性状态5、卫星DNA:主要分布在染色体着丝粒部位,由非常短的串联多次重复DNA序列组成,因为它的低复杂性又称简单序列DNA,又因其不同寻常的核苷酸组成,常在浮力密度离心中从整个基因组DNA中分离成一个或多个“卫星”条带,故称卫星DNA。
6、Southern印迹:将凝胶上分离的DNA片段转移到硝酸纤维素膜上,再通过同位素标记的单链DNA或RNA探针的杂交作用检测这些被转移的DNA 片段的方法。
步骤:限制性酶切DNA分子、琼脂糖凝胶电泳分离、碱变性、转膜、探针杂交、洗膜除去未杂交的探针、放射性自显影。
Nouthern印迹:将RNA分子从电泳凝胶转移到硝酸纤维素膜上,然后进行核酸杂交的一种那个实验方法。
Wouthren:将蛋白质从电泳凝胶中注意到硝酸纤维素膜上,然后与放射性同位素i125标记的特定蛋白质的抗体进行反应。
7、酶活力:指酶催化某化学反应的能力,其大小可以用在一定条件下所催化的某一化学反应的反应速率来表示,两者呈线性关系。
8、1)、可逆抑制作用:抑制剂与酶以非共价键结合,用透析、超滤或凝胶过滤等方法可以除去抑制剂,恢复酶活性。
主要包括:竞争性抑制、非竞争性抑制和反竞争性抑制作用三种。
竞争性抑制是I与S竞争E的结合部位,影响了S与E的正常结合。
非竞争抑制是I与S同时与E结合,但三元复合物不能进一步分解为产物,酶活性下降。
反竞争抑制是E只有与S结合后,才能与I结合,三元复合物不能进一步分解为产物。
2)、不可逆抑制作用:抑制剂通常以共价键与酶的必须基团进行不可逆结合,从而使酶失去活性。
酶的作用机制和酶的调节

别构激活剂 别构抑制剂
(2)别构酶的动力学
S形曲线(正协同) 表观双曲线(负协同效应)
(二)酶原的激活
酶原(zymogen):酶的无活性的前体
酶原的激活:由无活性的酶原转变为有活性 的酶的过程。酶原激活的实质是酶活性部位 的形成或暴露至分子的表面。
酶原激活的意义:在特定的环境和条件下发 挥作用;避免细胞自身消化;也可保证某些 特殊生理过程的正常进行,如凝血作用;有 的酶原可以视为酶的储存形式。
溶菌酶催化底物C1-O键裂解
五、酶活性的调节控制
(一)别构调控(allosteric regulation)
定义 别构调节:酶分子的非催化部位与某些
化合物可逆地非共价结合后发生构象的 改变,进而改变酶活性状态。 别构酶:具有别构现象的酶。 别构剂:能使酶分子发生别构作用的物
质。通常为小分子代谢物或辅因子
白
S
酶
SS
胰蛋白酶原
肠激酶
胰凝乳蛋白酶原
α-胰凝乳蛋白酶 +两个二肽
自
六肽
身 催
+
化
胰蛋白酶
弹性蛋白酶原 弹性蛋白酶 + 碎片
羧基肽酶原A
羧基肽酶A + 碎片
肠激酶启动的酶原激活
出血性胰腺炎发病机制?
凝血机制:1、受伤血管收缩减少血流;2、血小板粘聚成
栓堵住伤口;3、凝血相关因子的级联激活作用
蛋白激酶,磷酸化
酶
磷酸酶,脱磷酸化
酶-P
由核苷三磷酸(ATP)提供磷酸基,都需Mg2+。
酶的活性形式: 可能是磷酸化也可能是脱磷酸化
底物蛋白质被磷酸化的氨基酸残基有两类: (1)“P-O”键连接,如Thr, Ser, Tyr, Asp, Glu…… (2)“P-N”键连接,如Lys, Arg, His……
第四章 酶3 酶的作用机制及活性调节

• b. 修饰剂浓度与酶失活或降低的程度若成正比,则修饰位 于活性中心内
将修饰后的酶水解,肽键打开(但修饰剂与酶结合的共价 键不被打开)→得到带有标签的肽段→用氨基酸测序进行 鉴定
2、研究酶活性部位的方法
2、研究酶活性部位的方法
定点诱变法
• 改变编码蛋白质基因重的DNA顺序→改变氨基酸残基→确 定活性部位 • 如果被代换的氨基酸不影响酶的活性,则该位臵的氨基酸 残基不是必须基团 • 如果被代换的氨基酸使酶活性丧失或降低,则该位臵的原 有氨基酸残基是必须基团
• 1)vmax不变,Km值升高,该位臵氨基酸为结合基团
生
物
化
学
第四章 酶
§4.3 酶的作用机制和酶的调节
• 一、酶的活性部位 • 二、酶催化反应的独特性质 • 三、影响酶催化效率的有关因素
• 四、酶催化反应机制的实例
• 五、酶活性的调节控制
• 六、同工酶
一、酶的活性部位
• 只有少数的氨基酸残基参与底物结合及催化作用
• 酶的活性部位(active site/ active center )——与酶活力直 接相关的区域:分为结合部位(负责与底物的结合→决定酶 的专一性)和催化部位(负责催化底物键的断裂形成新键→ 决定酶的催化能力)
二、酶催化反应的独特性质
• 1、酶反应有两类:其一仅涉及到电子的转移(转换数约 108s-1);其二涉及到电子和质子两者或其他基团的转移 (约103s-1,大部分反应) • 2、酶催化作用是由氨基酸侧链上的功能基团(His、Lys、 Glu、Asp、Ser、Cys)和辅酶为媒介的→比只利用氨基酸 侧链来说,为催化反应提供了更多种类的功能基团 • 3、酶催化反应的最适pH范围通常是狭小的 • 4、与底物分子相比,酶分子很大而活性部位通常只比底物 稍大一些 • 5、存在一个或以上的催化基团及活性部位
第10章酶的作用机制和酶的调节

第10章酶的作用机制和酶的调节第10章酶的作用机制和酶的调节教学目的:掌握酶的活性部位结构与功能、酶活性的别构调节、酶原激活,了解酶高效性原因教学重点:酶活性部位的结构与功能及酶的活性的别构调节教学难点:酶活性的别构调节教学方法:多媒体教学内容:一、酶的活性部位及确定方法(一)酶活性部位概念及特点1、酶的活性中心(活性部位):指酶分子中的表面有一个必需基团比较集中、并构成一定空间结构的微小区域。
酶活性中心的基团,按其功能可分为结合基团和催化基团。
活性中心的基团都是维持酶活性的必需基团,2、酶活性部位的共同点:(1)酶活性部位仅占酶体积的很小一部分,通常只占整个酶分子体积的1~2%,酶分子是大分子物质,由很多氨基酸构成,而活性部位仅由几个氨基酸残基组成催化部位一般由2~3个氨基酸残基组成。
结合部位氨基酸残基数目,不同的酶有所不同。
可能是一个,也可能是多个。
(2)酶的活性部位具有三维结构,构成酶活性中心的基团,可位于同一条肽链上,也可位于不同的肽链上,在一级结构上可能相距甚远,但在空间结构上位置必须相互靠近;酶的空间结构受物理或化学因素影响时,酶的活性部位可能会遭破坏,酶会失活。
(3)活性中心的结合基团与底物专一性结合,这需要活性部位的基团精确排列。
活性部位具有一定的柔韧性,活性部位的结构并不是与底物的结构正好互补。
在酶与底物结合过程中,酶活性中心的构象在底物的诱导下可发生形变,然后嵌合互补形成中间产物,而底物在酶活性中心的诱导下也可发生形变,变的易与酶结合,有时是两者的构象同时发生变化后才互补契合(诱导契合学说)。
(4)酶活性部位位于酶分子表面的一个裂缝内,底物分子或底物分子的一部分结合到裂缝中,裂缝内的非极性基团较多,形成一个疏水环境,提高与底物的结合能力,也有极性的氨基酸残基,以便与底物结合并催化底物发生反应。
(5)底物通过较弱的次级键与酶结合。
组成酶活性中心的氨基酸残基,常见的有:组氨酸、赖氨酸、天冬氨酸、谷氨酸、丝氨酸、半胱氨酸和酪氨酸3、研究酶活性部位的方法(1)共价修饰(2)亲和标记法(3)切除法(4)X射线晶体结构分析法二、酶促反应机制(一)基元催化的分子机制:酶的催化作用包括若干基元催化。
第九章:酶的作用机制和酶的调节1

3.用于判断和确定酶活性中心的方法 1)酶的专一性研究 通过研究酶的专一性底物的结构特点,来判断和确定 活性中心的结构特点→确定活性中心的结构 研究酶的竞争性抑制剂的必需结构、酶与专一性底物 的相互关系,来确定酶活性中心结构。
2)酶分子侧链基团的化学修饰法 使用一些对酶分子侧链功能基团可进行共价修饰的 试剂作用与酶,以查出哪些基团是保持酶活力所必需 的。
三.与酶高效催化作用有关的因素 1.底物与酶邻近效应和定向效应 在酶促反应中,底物分子结合到酶的活性中心,一方 面底物在酶活性中心的有效浓度大大增加,有利于提高 反应速度; 另一方面,由于活性中心的立体结构和相关基团的诱 导和定向作用,使底物分子中参与反应的基团相互接近, 并被严格定向定位,使酶促反应具有高效率和专一性特 点。
第九章:酶的作用机制和酶的调节 1.酶的活性 部位 2.影响酶催 化效率的有关 因素 3.酶活性的 调节控制 4.同工酶
第一节:酶活性中心
以一个独立三级结构为完整生物共能分子最 高形式的酶,称为单体酶;以四级结构作为完整生物 功能分子结构的酶,称为寡聚酶。 1.酶的活性中心 酶蛋白中只有少数特定的氨基酸的侧链基团核 酶的催化活性直接有关,这些官能基团称为酶的必需 基团。在酶分子三级机构的构象中,由少数必需基团 组成的能与底物分子结合并完成特定催化反应的空间 小区域,称为酶的活性中心或酶活中心。构成酶活性 中心的必需基团,主要是某些氨基酸残基的侧链基团。
在酶的活性中心出现频率最高的氨基酸残基有:丝 氨酸、组氨酸、半胱氨酸、酪氨酸、天冬氨酸、谷氨酸 和赖氨酸,它们的极性侧链基团常常是酶活性中心的必 需基团。
2.酶的活性部位的特点
活性部位在酶分子的中提及中只占相当小的一 部分,通常只占整个酶分子体积的1%-2%。酶的活性 部位是一个三维实体 酶的底物部位并不是和底物的形状正好互补的, 而是在酶和底物结合的过程中,底物分子或酶分子, 有时是两者的构象发生了一定的变化后才互补的, 这时催化基团的位臵也正好在所催化底物键的断裂 和即将生成键的适当位臵。这个动态的辨认过程称 为诱导契合。 酶的活性部位是位于酶分子表面的一个裂缝内。 底物通过次级键较弱的力结合到酶上。 酶活性部位具有柔性或可运动性。
第六章 酶的作用机制和酶的调节 - 复制

5.酶除了具有进行催化反应必须的基团外, 还具有其他的特性
活性中心的实质
必需基团:活性 中心即酶分子中 在三维结构上相 互靠近的几个 aa残基或其上 的某些基团。 非必需基团:活 性中心以外的部 分对酶催化次要 但对活性中心形 成提供结构基础。
胰凝乳蛋白酶的活性中心
必须基团:酶表现催化活性不可缺少的基团
概念:指能催化相同的化学反应,但其结 构和理化性质及反应机理都有所不同的 一组酶。 应用: 在细胞分化及形态遗传的分子学基础研 究中很重要; 在代谢调控中起重要作用; 作为疾病诊断的指标。如乳酸脱氢酶 (LDH)
(四)酶原的激活:
1、概念: 酶原(proenzyme): 没有催化活性的酶的前体(precursor)。 酶原激活(活化): 从不具活性的酶原转变为有活性的酶的过 程。 其实质是一个或一些专一的肽被裂解, 使酶活性中心形成或暴露的过程。如:
结合部位:结合部位决定酶的专一性
结合部位
催化部位(Catalytic site)
催化 部位 决定 酶的 高效 性
三、影响酶催化效率的有关因素:
(一)底物与酶的邻近和定向效应:
S分子向E活性中心靠近,且趋向E催化部位, 使活性中心这一局部区域[S]增加,并使S分子 发生扭曲,易于断裂,降低反应所需活化能。 从而加快反应速度。
第六章 酶的作用机制和酶的调节
新疆农业大学农学院生物化学教研室 王希东 TEL:8763713 E-mail:wxdxnd@ wxd4085_cn@
本章主要内容
1. 酶的活性中心(活性部位) 2. 酶催化反应的独特性质 3. 影响酶催化效率的有关因素 4. 酶活性的调节
(一)别构酶 (allosteric enzymes):
第三节酶的作用机制和酶的调节

乳酸脱氢酶同工酶(LDHs)为四聚体,在体内共有
六种分子形式,即LDH1(H4),LDH2(H3M),LDH3 (H2M2),LDH4(HM3),LDH5(M4)和LDHx。
乳酸脱氢酶(LDH): M 三种类型的亚基 骨骼肌型
H x
心肌型
x亚基
亚基 组成
H H H H LDH1 (H4)
M H H H LDH2 (M H3)
7.多功能催化作用
酶的活性中心部位,一般都含有多个起催 化作用的基团,这些基团在空间有特殊的 排列和取向,可以对底物价键的形变和极 化及调整底物基团的位置等起到协同作用, 从而使底物达到最佳反应状态。
三、酶活性的调节控制
生物体内的各种生理活动均以一定的物 质代谢为基础。为了适应某种生理活动的 变化,需要对一定的代谢活动进行调节。 通过对酶的催化活性的调节,就可以达 到调节代谢活动的目的。 可以通过改变其催化活性而使整个代谢反 应的速度或方向发生改变的酶就称为限速 酶或关键酶。
胰腺的蛋白酶
胰腺中存在胰蛋白酶、胰凝乳蛋白酶 和弹性蛋白酶的酶 原,它们转移到小肠后,首先是胰蛋白酶原被肠激酶断 裂,然后胰蛋白酶进一步激活其它两个酶。 胰蛋白酶原 肠激酶
胰蛋白酶
胰凝乳蛋白酶原 弹性蛋白酶原 弹性蛋白酶
胰凝乳蛋白酶
胰凝乳蛋白酶的活化过程
胰凝乳蛋白酶原(无活性)
1 13 14 15 16 146147148149 245
5.金属离子的催化作用
金属离子的催化作用往往和酸的催化作用相似,有多 种途径参加催化过程。(结合底物为反应定向;调节 氧化还原反应;静电稳定或屏蔽负电荷)
6.微环境的影响(酶活性中心是低介电区域)
酶活性中心处于一个非极性环境中, 从而有利于同底物的结合。 (水的极性和形成氢键的能力都较强, 能够减弱极性基团间的相互作用。)
酶活性调节

6、 多元催化和协同催化 酶催化反应的高效性,可能是由于以上五种因素中的几种因 素协同作用的结果,而非酶催化反应往往只有一种催化机制。
7、 活性部位微环境的影响 疏水环境 介电常数低,加强了极性基团间的作用。
三、 酶活性的别构调节
别构酶:一般都是寡聚酶,通过次级键由多亚基构 成,这种酶除了有活性中心外,还有一个别构中 心,当调节物结合到别构中心上时,会引起酶分 子构象发生变化而导致酶活性的变化。 别构调节:调节物(效应物)与别构酶分子中的 别构中心(调节中心)非共价结合后,酶分子构 象发生改变,从而调节酶的活性,即别构调节。
别构调节举例:
天冬氨酸转氨甲酰酶(ATCase)是一个研究的比较透明的别构酶。它催 化下列反应:
这个酶是合成CTP的多酶体系反应序列中的第一个酶,反应系列的终产物CTP是该 酶的别构抑制剂,而ATP则是它的别构激活剂。 ATCase由12条多肽链组成,其中6条形成2个三聚体称催化亚基(每个三聚体 有3个结合天冬氨酸的底物结合部位),6条形成3个二聚体称调节亚基( 每个二聚 体有2 个别构效应剂CTP或ATP的结合部位。见下图解:
5、 别构酶的鉴定
双倒数作图不是直线
谢谢 大 家
二、 影响酶催化效率的有关因素
1、 酸碱催化 酶分子的一些功能基团起瞬时质子供体或质子受体的作用。 分为狭义的酸碱催化和广义的酸碱催化。 酶分子中可作为广义酸碱的功能基团:氨基、羧基、巯基、 酚羟基、咪唑基。
影响酸碱催化反应速度的两个因素: ⑴酸碱强度(pK值)。组氨酸咪唑基的解离常数为6,在pH6附 近给出质子和结合质子能力相同,是最活泼的催化基团。 ⑵给出质子或结合质子的速度。咪唑基最快,半寿期小于10-10 秒。Leabharlann 2、 共价催化
酶

酶作用专一性的机制
结构性质互补假说(Structure-properties complementation theory)
底物的结构和酶活性中心三维空间结构互补外,在酶和底物的性质方面也有要求。 如果底物带电荷,酶的活性中心必带相反电荷,同时底物和活性中心的极性也必 然相同。
酶作用高效率的机制
Cleft,crevice or cavities(裂缝、凹穴、裂沟)为疏水的微环境
(二)酶活性中心的结构特点
1.活性中心只占酶分子总体积的很小一部分,往往只占整个 酶分子体积的1%-2%。 2.酶的活性部位是一个三维实体,具有三维空间结构。活性 中心的空间构象不是刚性的,在与底物接触时表 现出一定的柔性和运动性。 3.酶的活性部位并不是和底物的形状正好互补的,而 是在酶和底物的结合过程中,底物分子或酶分子、有时是 两者的构象同时发生了一定的变化后才互补的,此时催化 基团的位置正好处在所催化底物键的断裂和即将生成键的 适当位置,这个动态辨认过程称为诱导契合(induced-fit).
酶-底复合物形成时,酶分子构象发生变化,底物分子 也常常受到酶的作用而发生变化,甚至使底物分子发 生扭曲变形,从而使底物分子某些键的键能减弱,产 生键扭曲,有助于过度态的中间产物形成,从而降低了 反应的活化能。
(四) 多功能催化作用
酶的活性中心部位,一般都含有多个起催化作用的 基团,这些基团在空间有特殊的排列和取向,可以 对底物价键的形变和极化及调整底物基团的位置等 起到协同作用,从而使底物达到最佳反应状态。
01 酶的活性中心
02 酶作用专一性机制 03 酶作用高效率的机制
01
(一)基本概念: 酶的活性中心是指结合底物和将底物转化为产物的区 域,通常是相隔很远的氨基酸残基形成的三维实体。 酶的活性中心包括两个功能部位:结合部位和催化部 位。
酶的作用机制和酶的活性部位调节

金属离子的作用: 作为亲电催化剂稳定反应时形成的负电荷,以利于底物进
入反应过渡态。
(二)酶具有高催化能力的原因
1. 邻近效应与定向效应 作用:使分子间的反应变成类似分子内的反应
邻近效应(proximity effect) :中间复合物的形成使有效 浓度极大升高。
接触残基:R1、R2、R6、 R8、R9、R163
辅助残基:R3、R4、R5、 R164、R165
结构残基:R10、R162、 R169
非贡献残基:其它
2. 酶活性部位的一般特点
① 只 占 酶 整 体 的 相 当 小 的 一 部 分 ( 1%~2% ) 。
② 是一个三维实体。
③ 底物结合的专一性决定于活性部位中精确地 原子排列,即直接契合或诱导契合。
酶活性部位:结合部位+催化部位
结合基团(binding group):参与和底物结合的必需基团; 决定酶的专一性。
催化基团(catalytic group):催化使底物转变为产物的 化学反应的必需基团;决定酶的催化能力。
必需基团:
某些氨基酸残基的侧链、有时也 包括辅酶分子或它的基团。
酶蛋白
异促别构效应
(heterotropic allosteric effect)
必需残基 非必需残基
活性中心
结合残基 接触残基
催化残基
辅助残基
活性中心外 非贡献残基
结构残基
酶分子中的残基分为四类:
接触残基:负责底物的结合与催化 辅助残基:起协助作用; 结构残基:维持酶的构象; 非贡献残基:它的替换对活性无影响,但对酶的
免疫、运输、调控与寿命等有作用。பைடு நூலகம்前二者构成活性中心,前三者称为酶的必需基团。
酶的活性部位
的羟基,半胱氨酸的 H2N CH C OH
N
巯基和组氨酸的咪唑
CH2
N
基。
H
N
NH
O
H2N CH C OH O
CH2 H2N CH C OH
COOH
酸、碱性基团:
CH2 CO
CH2 CO
天冬氨酸和谷氨
OH
O OH
酸的羧基,酪氨 O H2N CH C OH源自NH 2酸的酚羟基
H2N CH C OH CH2
广义酸-碱催化是指通过质子酸提供部分质子, 或是通过质子碱接受部分质子的作用,达到降 低反应活化能的过程。
酶分子中可以作为广义酸、碱的基团
广义酸基团 (质子供体)
+
-COOH, -NH 3, -SH,
广义碱基团
(质子受体)
-COO -,
..
-NH 2,
-S ,-
+
OH HN NH
O- :N NH
结合部位决定酶的专一性, 催化部位决定酶所催化反应
的性质。
补:酶活性中心的必需基团
顺便复习一些重要AA的R基 O
H2N CH C OH
OH
酶必需基团指酶活性
CH2
中心实现催化作用的
OH O
必需的些氨基酸基团。 H2N CH C OH
SH
主要包括:
CH2
亲核性基团:丝氨酸
O SH
第十章 酶的催化作用 机制和酶的调节
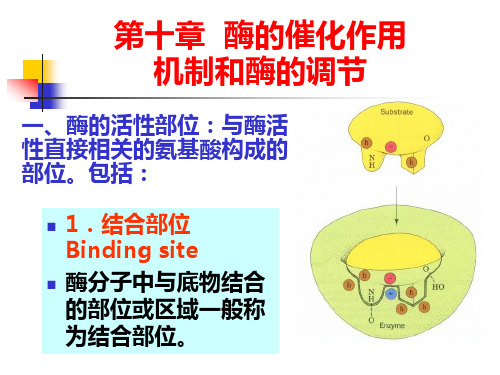
一、酶的活性部位:与酶活 性直接相关的氨基酸构成的 部位。包括:
1.结合部位 Binding site
酶分子中与底物结合 的部位或区域一般称 为结合部位。
2.催化部位 catalytic site
酶的作用机制和酶的活性部位调节.ppt

上述加速酶促反应的诸因素,不是同时在一种酶中起作用。 不同的酶,起作用的主要因素是不一样的,每种酶都有自己
的特点。
三、酶活性的别构调节
酶的调节
酶活性的调节(细调)
–别构调节 –酶原激活 –共价调节
酶数量的调节(粗调)
控制酶的合成和降解速度
(一) 酶的别构效应和别构酶
第11章 酶作用机制和酶活性调节
主要内容
酶的活性部位 酶促反应机制 酶活性的别构调节 酶活性的共价调节 同工酶
一、酶的活性部位
1.酶的活性部位(active site,活性中心active center):
是指酶分子的表面有一个必需基团比较集中、并构成一定的空 间结构的微小区域,在这里必需基团参与和底物结合并完成 把底物转变成产物的化学反应。
暂时性地向底物提供质子或从底物接纳质子以稳定过渡态 的一种催化机制。
特殊的酸碱催化 (或狭义) :
一般的酸碱催化
√ (或广义) :
H+和OH-的催化作用
质子酸提供质子,或是质子碱 接受质子的作用。
2.共价催化(covalent catalysis)
催化剂通过与底物形成反应活性很高的共价过渡产物,使 反应活化能降低,从而提高反应速度的过程。 包括亲核催化(为主)和亲电催化。
④ 大多数底物都是以相当弱的力与酶活性部位 结合的,这些力与稳定酶(蛋白质)三维结 构的力基本相同。
⑤ 活性部位是酶分子表面的一个空穴或裂沟。 疏水的微环境下有利于极性氨基酸残基发生 结合和催化。
频率最高的活性中心的氨基酸残基: Ser、His、Cys、Tyr、Asp、Glu、Lys。
Ser
Glu
别构效应(allosteric effect):
酶的活性调节

(二)E 的活性中心特点 1 几个氨基酸残基,1%〜2 %酶分子体积
384
(二) E 的活性中心特点
2 3
三维实体 表面或接近表面
裂缝(crevice)
疏水区域
4 柔性或可运动性
E 诱导契合和 S底物的形变
5
ES 是由次级键形成
384
酶的活性中心示意图
酶的结构
活性中心
必需基团
结合部位 催化部位 活性中心以外的必需基团
长的凹穴。最适底物正好与
酶分子的凹穴相结合,凹穴
中的Glu35和Asp52 是活性中 心的氨基酸残基。
2. 催化作用机理 • 溶菌酶底物与酶活性中心的关系
溶菌酶活性中心上的Asp52氧 原子距离底物敏感键(C-O键)中 碳原子只有0.3nm,活性中心 上另一个氨基酸 Glu35的羧基 距离底物敏感键(C-O键)中氧原 子也只有0.3nm,溶菌酶的活 性中心的氨基酸残基与底物敏 感键既靠近又定向。
接有关,即与酶活力直接相关的区域称为酶的活性部位。
酶的活性部位是酶分子进行催化反应的一个场所,是酶分子的一小 部分区域,在这个区域上的少数几个特异的氨基酸参与结合底物催化底 物,把酶分子上的这个区域称为酶的活性部位。
结合部位
负责酶与底物的结合,决定
活性 部位
催化部位
酶 的专一性
负责催化底物,决定酶
酶活性中心的羧基与水形成氢键,导 致酶活性中心羧基表面有一层水化层,水 分子的屏蔽作用,大大削弱了酶分子与底 物离子间的静电相互引力,不利于酶促反 应。
酶催化作用机理: 综上所述:
酶与底物结合时,由于酶的变形(诱导契合) 或底物变形使二者相互适合,并依靠离子键、氢 键、范德华力的作用和水的影响,结合成中间产 物,在酶分子的非极性区域内,由于酶与底物的 邻近、定向,使二者可以通过亲核\亲电催化、
第11章 酶作用机制和酶活性调节

胰蛋白酶激活系统
2)凝血系统蛋白酶原的激活
四、同工酶
同工酶:能催化相同的化学反应,但其蛋白质分子结构、理化
性质和免疫性能等方面都存在明显差异的一组酶。
乳酸脱氢酶(LDH)催化的反应
CH3 C O + NADH + H COOH +
LDH
CH3 CHOH + NAD COOH +
不同组织中LDH同工酶的电泳图谱
逆的非共价结合,使酶构象改变,从而改变酶的催化活性的现 象。 正别构效应:因别构导致酶活性增加 别构效应 负别构效应:因别构导致酶活性降低
具有别构效应的酶,也称变构酶(allosteric enzyme)。
变构抑制效应
2、变构调节的机制
变构效应剂 + 酶的调节亚基 疏松或紧密 亚基聚合或亚基解聚 形成酶分子多聚体
(2)一般由相距较远的2-3个氨基酸残基组成
(3)通过诱导契合和底物形变实现酶和底物的结合
诱导契合:酶与底物相互接近时,酶在底物的诱导下发生构象变 化的过程。这一学说称为诱导契合假说(induced-fit hypothesis)。 底物形变:当酶与一旦底物结合,酶就能使底物发生形变。其本 质是酶中某些基团使底物分子中某些基团的电子云密度重新分配而 产生电子张力,造成底物发生形变,使底物更加接近它的过渡态, 促使旧键弱化新键形成,使反应易于发生。
(4)底物通过次级键结合到酶上
(5)底物通过次级键结合到酶上
二)酶活性中心的鉴定方法
1、切除法 对小分子且结构已知的酶多用此法。用专一性的
酶切除一段肽链后剩余的肽链仍有活性,说明切除的
肽链与活性无关,反之,切除的肽链与活性有关。
2、化学修饰法 选用适当的化学试剂与酶蛋白中的氨基酸残 基的侧链基团发生反应引起共价结合、氧化或还原
酶的活性中心包括( )和( )两部分

酶的活性中心包括( )和( )两部分
酶活性中心有一个结合部位和一个催化部位,分别决定专一性和催化效率,是酶分子发挥作用的一个关键性小区域。
一般认为活性中心主要由两个功能部位组成:第一个是结合部位,酶的底物靠此部位结合到酶分子上;第二个是催化部位,底物的键在此被打断或形成新的键从而发生一定的化学变化。
组成功能部位的是酶分子中在三维结构上比较靠近的少数几个氨基酸残基或是这些残
基上的某些基团,它们在一级结构上可能相距甚远,甚至位于不同肽链上,而是通过肽链的盘绕、折叠在空间构象上相互靠近;对于需要辅酶的酶来说,辅酶分子或辅酶分子的某一部分结构也是功能部位的组成部分。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
OH
酶必需基团指酶活性
CH2
中心实现催化作用的
OH O
必需的些氨基酸基团。 H2N CH C OH
SH
主要包括:
CH2
亲核性基团:丝氨酸
O SH
的羟基,半胱氨酸的 H2N CH C OH
N
巯基和组氨酸的咪唑
CH2
N
基。
H
N
NH
O
H2N CH C OH O
CH2 H2N CH C OH
COOH
2.催化部位 catalytic site
酶分子中促使底物发生化学 变化的部位称为催化部位。
通常将酶的结合部位和催化 部位总称为酶的活性部位或 活性中心。
结合部位决定酶的专一性, 催化部位决定酶所催化反应
的性质。
补:酶活性中心的必需基团
顺便复习一些重要AA的R基 O
H2N CH C OH
形变的机制:与酶结合,底物分子内敏感键中电子云 密度增高或降低,使敏感键更敏感,从而底物分子形 变。
3、 酸碱催化
指通过瞬时的向反应物提供质子或从反应物接 受质子以稳定过渡态,加速反应的催化机制。
酸-碱催化可分为狭义的酸-碱催化和广义的酸 -碱催化。酶参与的酸-碱催化反应一般都是广 义的酸-碱催化方式。
O CH3 C O
..
N
NH
O CO
..
N
NO2
CH3
O C
N+
NH
+
O-
O
NO2
+
N
+
O-
NO2
NO2
NH
NH
2、底物形变和诱导契合
该学说认为酶表面并没有一种与底物互补的固定形状, 而只是由于底物的诱导才形成了互补形状,同时底物 了发生形变,接近其过渡态,降低了反应活化能,使 反应易于发生.
1958年,他参加发起人工合成胰岛素工作,并负责胰岛素 A和B链的拆合。
邹承鲁等用自己创立的动力学方法比较研究了各种不同类型 的酶在变性过程中构象和活力变化的关系,得到活性丧失先 于构象变化的结果,据此提出了酶活性部位柔性的假说。
——邹承鲁
邹承鲁:我的学生差不多都退休了。 学生:就先生还在这干。我们也快退休了,但是老
先生还得干。 邹承鲁:我是退休嫌闷得慌。 学生:其实搞科学的就是这样,他也不求什么,就
求一个能干自己想干的事的环境。
酶催化作用的本质
酶催化作用的本质是酶的活性中心与底 物分子通过短程非共价力(如氢键,离子 键和疏水键等)的作用,形成E-S反应中 间物。
其结果使底物的价键状态发生形变或极 化,起到激活底物分子和降低过渡态活 化能作用。
广义酸-碱催化是指通过质子酸提供部分质子, 或是通过质子碱接受部分质子的作用,达到降 低反应活化能的过程。
酶分子中可以作为广义酸、碱的基团
广义酸基团 (质子供体)
+
-COOH, -NH 3, -SH,
广义碱基团
(质子受体)
-COO -,
..
-NH 2,
-S ,-
+
OH HN NH
O- :N NH
邹承鲁(1923年5月17日—),出生于山东青岛,祖籍江苏 无锡。生物化学家。1980年当选为中国科学院学部委员(院 士)。
1946年,赴英国剑桥大学。研究生期间,他在国际上最早 用蛋白水解酶部分水解方法研究蛋白质结构与功能的关系, 单独署名的论文在英国《自然》杂志发表。1951年,邹承 鲁获英国剑桥大学生物化学博士学位。
以3种主要途径参加催化过程: (1)通过结合底物为反应定向
(2)通过可逆改变金属离子 氧化态调节氧化还原反应
许多氧化-还原酶 中都含有铜或铁离 子,它们作为酶的 辅助因子起着传递 电子的功能。
Fe3+ e Fe2+
酶活性部位的共同特点
(1)活性部位酶占总体很小部分:1-2% (2)酶活性部位是一个三维实体 (3)酶活性部位与底物通过诱导契合后
结合 (4)酶活性部位是其表面一个裂缝,内
部非极性基团较多,是一疏水区域 (5)底物通过非共价与酶结合 (6)活性部位具有柔性或可运动性
著名生物化学家、中科院院士
*二、酶催化反应的独特性质 (自学,以增对酶催化作用所理解)
三、影响酶催化效率的有关因素
酶是专一性强, 催化效率很高的 生物催化剂,这 是由酶分子的特 殊结构决定的。 多种因素可以使 酶催化的反应加 速:
1、底物和酶的邻近效应和定向效应
邻近效应:酶与底物形成复合物后,使底物和 底物之间(对多底物反应而言),酶的催化基 团与底物之间结合于同一分子使反应有效浓度 大提高,有利于提高反应速率大增加。
His 是酶的酸碱催化作用中最活泼的一个 催化功能团。
4、 共价催化
基本原理: 又称亲核催化或亲电子催化,催化
时,酶分子上亲核基团或亲电子基 团分别放出电子或汲取电子并作用 于底物的缺电子或富电子中心,迅 速形成不稳定的共价中间复合物, 降低反应活化能,使反应加速。 酶上常见的亲核基团: Ser 的羟 基, Cys 的巯基, His 的咪唑基。 底物上亲电基团:磷酰基,酰基, 糖基。 ——P392实例
5、 金属离子催化作用
*根据金属离子与酶蛋白结合程度,可分为两类: 金属酶和金属—激活酶。 (1)金属酶:酶蛋白与金属离子结合紧密。如 Fe2+/ Fe3+ 、Cu+/Cu3、Zn2+ 、Mn2+、Co2 等过 渡态离子。 (2)金属—激活酶:金属离子与酶的结合一般 较松散。在溶液中,酶与这类离子结合而被激 活。如Na+ 、K+、 Mg2+、 Ca2+CO
天冬氨酸和谷氨
OH
O OH
酸的羧基,酪氨 O H2N CH C OH
NH 2
酸的酚羟基
H2N CH C OH CH2
CH2
赖氨酸的氨基,
CH2
OH
组氨酸的咪唑基。 CH2
CH2
NH 2 OH
补:酶的调节部位 Regulatory site
某些酶分子中存在着一些可以与其他分子发生 某种程度的结合的部位,从而引起酶分子空间 构象的变化,对酶起激活或抑制作用。
定向效应:反应物的反应基团之间和酶的催化 基团与底物的反应基团之间正确取位,使底物 分子中参与反应的基团相互接近,并被严格定 定位,使酶促反应具有高效率和专一性特点。 (例如:P389邻苯丙酸内酯形成的反应)
例:邻近效应
以乙酸对硝基苯酯以咪唑作为酶催化的水解反应为例, 若将咪唑连到其上,则反应加快24倍