生物合成详解演示文稿
合集下载
《生物合成PHA》PPT课件

– 当限制性营养物再次被提供时,PHAs能被胞内酶降解 后作为碳源和能源利用。
17
精选PPT
17
• 胞中积累的PHAs存在形式
– 以单个粒子的形态存在,每个细胞含有的颗粒 数量的大小随微生物种类而不同,在Ralstonia eutropha中,每个细胞含有8-10个颗粒,每个 颗粒直径大小为0.2-0.5μm;
• 由于PHAs具有低溶解性和高分子量,它在胞内的积累不 会引起渗透压的增加,是理想的胞内贮藏物,比糖原、 多聚磷酸或脂肪更加普遍地存在于微生物中。
• PHAs的通式可写成:
R
O
单体数目
___
O
__
CH__
CH2__
C
___ n
5
精选PPT
5
聚合物命名
• R为甲基时,其聚合物为聚β--羟基丁酸(PHB) • R为乙基时,其聚合物为聚β--羟基戊酸(PHV) • 在一定条件下两种或两种以上的单体还能形成共聚物,其
• 目前研究的较多的微生物: – 产碱杆菌属(Alcaligenes europhus, 现在更名为Ralstonia eutropha) – 假单胞菌属(Pseudonomas) – 甲基营养菌(Methylotrophs) – 固氮菌属(Azotobacter) – 红螺菌属(Rhodospirilum)
3
其中,m=1,2 或3,n 为单体数目,R 代表侧链,多
为C1~C13 的不同链长的正烷基,也可以是支链,不饱
和的或带取代基的烷基。
当m=1 时,R 为甲基时,为
(PHB),
是PHA中最常见而又最重要的一种。
精选PPT
4
二、PHAs的结构、物理化学性质和应用
17
精选PPT
17
• 胞中积累的PHAs存在形式
– 以单个粒子的形态存在,每个细胞含有的颗粒 数量的大小随微生物种类而不同,在Ralstonia eutropha中,每个细胞含有8-10个颗粒,每个 颗粒直径大小为0.2-0.5μm;
• 由于PHAs具有低溶解性和高分子量,它在胞内的积累不 会引起渗透压的增加,是理想的胞内贮藏物,比糖原、 多聚磷酸或脂肪更加普遍地存在于微生物中。
• PHAs的通式可写成:
R
O
单体数目
___
O
__
CH__
CH2__
C
___ n
5
精选PPT
5
聚合物命名
• R为甲基时,其聚合物为聚β--羟基丁酸(PHB) • R为乙基时,其聚合物为聚β--羟基戊酸(PHV) • 在一定条件下两种或两种以上的单体还能形成共聚物,其
• 目前研究的较多的微生物: – 产碱杆菌属(Alcaligenes europhus, 现在更名为Ralstonia eutropha) – 假单胞菌属(Pseudonomas) – 甲基营养菌(Methylotrophs) – 固氮菌属(Azotobacter) – 红螺菌属(Rhodospirilum)
3
其中,m=1,2 或3,n 为单体数目,R 代表侧链,多
为C1~C13 的不同链长的正烷基,也可以是支链,不饱
和的或带取代基的烷基。
当m=1 时,R 为甲基时,为
(PHB),
是PHA中最常见而又最重要的一种。
精选PPT
4
二、PHAs的结构、物理化学性质和应用
生物合成反应PPT参考幻灯片

MeO
OH
左炔诺孕酮
O
24
氧化反应
+ + + Sub
NAD(P)H
H+
单加氧酶 O2
+ + SubO
NAD(P) +
H2O
辅酶循环
+ Sub
O2
双加氧酶
SubO2
+ O2
2e-
氧化酶
2H+ O2 2-
H2O2
氧化酶
4H+
+ O2
2e-
2O 2-
H2O
25
单加氧酶催化的氧化反应(细胞色素P450单加氧酶和黄
O S
COCl
S
CO2H N
H L-脯 氨 酸
CH3OH,NaOH
HO2C
S
SC N
SH
O
皱落假丝酵母
1) SOCl2
CO2H
S CO2H 2)
CO2H
OH
N
H
NaHS
HO2C
S
SC N
Cl
O
46
Cl CHO
Cl
CH2(COOEt)2 哌啶,EtOH
CH
(EtO2C)2HC
CH(CO2Et)2
Cl CH(CH2COOH)2
马肝醇HL脱ADH氢酶
布氏T热BA厌DH氧菌醇脱氢酶
羟基H甾SDH体脱氢酶
常用脱氢酶底物优先结构特征
11
酵母细胞(Yeast)的还原反应
简单无环酮的还原与脱氢酶相似,遵循Prelog规则
O
R1
R2
面包酵母
H OH
R1
DNA的生物合成课件

目录
目录
目录
目录
AT GC GC TA AT CG TA GC CG CG AT CG TA GC GC
母链DNA
CG CG AT CG TA GC GC
复制过程中形成 的复制叉
AT
AT
GC
GC
GC
GC
TA
TA
AT
AT
CG
CG
TA
TA
GC + GC
CG
CG
CG
CG
AT
AT
CG
CG
TA
TA
GC
功能 • 维持染色体的稳定性
• 维持DNA复制的完整性
70
目录
•端粒酶(telomerase)
组成
• 端粒酶RNA (human telomerase RNA, hTR) • 端粒酶协同蛋白(human telomerase associated
protein 1, hTP1) • 端粒酶逆转录酶(human telomerase reverse
46
目录
一、原核生物的DNA生物合成
(一)复制的起始
需要解决两个问题: 1. DNA解开成单链,提供模板。 2. 合成引物,提供3-OH末端。
47
目录
1. DNA解链
1
13 17
29 32
44
GATTNTTTATTT · · · GATCTNTTNTATT · · · GATCTCTTATTAG · · ·
43
目录
5’
O
3’
O P O-
O-
DNA连接酶
HO
ATP ADP
5’
O
3’
目录
目录
目录
AT GC GC TA AT CG TA GC CG CG AT CG TA GC GC
母链DNA
CG CG AT CG TA GC GC
复制过程中形成 的复制叉
AT
AT
GC
GC
GC
GC
TA
TA
AT
AT
CG
CG
TA
TA
GC + GC
CG
CG
CG
CG
AT
AT
CG
CG
TA
TA
GC
功能 • 维持染色体的稳定性
• 维持DNA复制的完整性
70
目录
•端粒酶(telomerase)
组成
• 端粒酶RNA (human telomerase RNA, hTR) • 端粒酶协同蛋白(human telomerase associated
protein 1, hTP1) • 端粒酶逆转录酶(human telomerase reverse
46
目录
一、原核生物的DNA生物合成
(一)复制的起始
需要解决两个问题: 1. DNA解开成单链,提供模板。 2. 合成引物,提供3-OH末端。
47
目录
1. DNA解链
1
13 17
29 32
44
GATTNTTTATTT · · · GATCTNTTNTATT · · · GATCTCTTATTAG · · ·
43
目录
5’
O
3’
O P O-
O-
DNA连接酶
HO
ATP ADP
5’
O
3’
第十章蛋白质的生物合成PPT课件
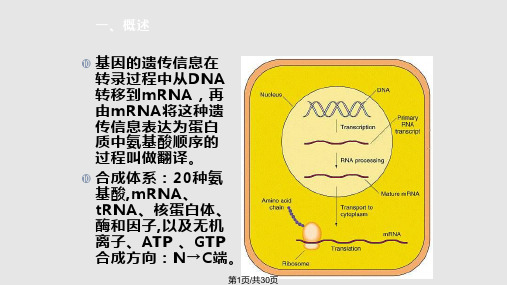
第15页/共30页
肽链的延长
进位 (氨酰tRNA进入A位点) 参与因子:延长因子EFTu(Tu)、EFTs(Ts)、GTP、氨酰tRNA 肽链的形成 肽酰基从P位点转移到A位点,形成新的肽链 移位(translocase) 在移位因子(移位酶)EF-G的作用下,核糖体沿mRNA(5’-3’) 作相对移动,使原来在A位点的肽酰-tRNA回到P位点
第2页/共30页
第3页/共30页
原核生物 核糖体组 成
真核生物 核糖体组 成
第4页/共30页
2. tRNA
结合氨基酸:一种氨基 酸有几种tRNA携带, 结合需要ATP供能,氨 基酸结合在tRNA3‘CCA的位置。
反密码子:每种tRNA的 反密码子,决定了所带 氨基酸能准确的在 mRNA上对号入座 。 反密码子与mRNA的第 三个核苷酸配对时,不 严格遵从碱基配对原则
三、遗传密码子
• 为一个氨基酸编码进入蛋白质多肽链特定线性 位置的三个核苷酸单位称为密码子(Coden) 或三联体密码。
• 密码子的发现
• 统计学方法 • 人工合成仅由一种核苷酸组成的多聚核苷酸,推测由
哪一种氨基酸合成的多肽 • 核糖体结合试验 1965年,Nirenberg用poly u加入
C14标记的20种aa,仅有苯丙氨酸的寡肽,UUU=苯丙 氨酸,用此法破译了全部密码,编出遗传密码表。
第22页/共30页
五、真核细胞蛋白质合成的特点
核糖体为80S,由60S的大亚基和40S的小亚基组成 起始密码AUG 起始tRNA为Met-tRNA 起始复合物结合在mRNA 5’端AUG上游的帽子结构,真核mRNA无富含嘌
呤的SD序列(除某些病毒mRNA外) 已发现的真核起始因子有近9种(eukaryote Initiation factor,eIF) 复合物称
肽链的延长
进位 (氨酰tRNA进入A位点) 参与因子:延长因子EFTu(Tu)、EFTs(Ts)、GTP、氨酰tRNA 肽链的形成 肽酰基从P位点转移到A位点,形成新的肽链 移位(translocase) 在移位因子(移位酶)EF-G的作用下,核糖体沿mRNA(5’-3’) 作相对移动,使原来在A位点的肽酰-tRNA回到P位点
第2页/共30页
第3页/共30页
原核生物 核糖体组 成
真核生物 核糖体组 成
第4页/共30页
2. tRNA
结合氨基酸:一种氨基 酸有几种tRNA携带, 结合需要ATP供能,氨 基酸结合在tRNA3‘CCA的位置。
反密码子:每种tRNA的 反密码子,决定了所带 氨基酸能准确的在 mRNA上对号入座 。 反密码子与mRNA的第 三个核苷酸配对时,不 严格遵从碱基配对原则
三、遗传密码子
• 为一个氨基酸编码进入蛋白质多肽链特定线性 位置的三个核苷酸单位称为密码子(Coden) 或三联体密码。
• 密码子的发现
• 统计学方法 • 人工合成仅由一种核苷酸组成的多聚核苷酸,推测由
哪一种氨基酸合成的多肽 • 核糖体结合试验 1965年,Nirenberg用poly u加入
C14标记的20种aa,仅有苯丙氨酸的寡肽,UUU=苯丙 氨酸,用此法破译了全部密码,编出遗传密码表。
第22页/共30页
五、真核细胞蛋白质合成的特点
核糖体为80S,由60S的大亚基和40S的小亚基组成 起始密码AUG 起始tRNA为Met-tRNA 起始复合物结合在mRNA 5’端AUG上游的帽子结构,真核mRNA无富含嘌
呤的SD序列(除某些病毒mRNA外) 已发现的真核起始因子有近9种(eukaryote Initiation factor,eIF) 复合物称
蛋白质生物合成PPT课件演示教学.ppt

缬 脯 苏 天冬
缬 丙 酪 甘
缬 丙 丝 精
3. 简并性(degeneracy)
1. 核糖体大小亚基分离; 2. 核糖体小亚基结合于mRNA的起始密码子附近; 3. fMet-tRNAfMet结合在核糖体P位 ; 4. 核糖体大亚基结合形成起始复合物。
一、翻译起始复合物的装配启动肽链合成
(a)起始复合物的装配过程;(b)rRNA识别mRNA的核糖体结合位点,保证翻译起始在起始密码子处
密码子(codon)
起始密码子和终止密码子:
遗传密码表
遗传密码的特点
1. 方向性(directional)
翻译时遗传密码的阅读方向是5→3,即读码从mRNA的起始密码子AUG开始,按5→3的方向逐一阅读,直至终止密码子。
N
C
肽链延伸方向
5
3
读码方向
2. 连续性(non-punctuated)
23S-rRNA 5S-rRNA
18S-rRNA
28S-rRNA 5.8S-rRNA 5S-rRNA
蛋白质
rpS 21种
rpL 36种
rpS 33种
rpL 49种
不同细胞核蛋白体的组成
核蛋白体的组成
核糖体在翻译中的功能部位
四、肽链生物合成需要酶类和 蛋白质因子
氨基酰-tRNA合成酶(aminoacyltRNA synthetase),催化氨基酸的活化; 转肽酶(peptidase),催化核蛋白体P位上的肽酰基转移至A位氨基酰-tRNA的氨基上,使酰基与氨基结合形成肽键;并受释放因子的作用后发生变构,表现出酯酶的水解活性,使P位上的肽链与tRNA分离; 转位酶(translocase),催化核蛋白体向mRNA3’-端移动一个密码子的距离,使下一个密码子定位于A位。
缬 丙 酪 甘
缬 丙 丝 精
3. 简并性(degeneracy)
1. 核糖体大小亚基分离; 2. 核糖体小亚基结合于mRNA的起始密码子附近; 3. fMet-tRNAfMet结合在核糖体P位 ; 4. 核糖体大亚基结合形成起始复合物。
一、翻译起始复合物的装配启动肽链合成
(a)起始复合物的装配过程;(b)rRNA识别mRNA的核糖体结合位点,保证翻译起始在起始密码子处
密码子(codon)
起始密码子和终止密码子:
遗传密码表
遗传密码的特点
1. 方向性(directional)
翻译时遗传密码的阅读方向是5→3,即读码从mRNA的起始密码子AUG开始,按5→3的方向逐一阅读,直至终止密码子。
N
C
肽链延伸方向
5
3
读码方向
2. 连续性(non-punctuated)
23S-rRNA 5S-rRNA
18S-rRNA
28S-rRNA 5.8S-rRNA 5S-rRNA
蛋白质
rpS 21种
rpL 36种
rpS 33种
rpL 49种
不同细胞核蛋白体的组成
核蛋白体的组成
核糖体在翻译中的功能部位
四、肽链生物合成需要酶类和 蛋白质因子
氨基酰-tRNA合成酶(aminoacyltRNA synthetase),催化氨基酸的活化; 转肽酶(peptidase),催化核蛋白体P位上的肽酰基转移至A位氨基酰-tRNA的氨基上,使酰基与氨基结合形成肽键;并受释放因子的作用后发生变构,表现出酯酶的水解活性,使P位上的肽链与tRNA分离; 转位酶(translocase),催化核蛋白体向mRNA3’-端移动一个密码子的距离,使下一个密码子定位于A位。
脂肪酸的生物合成及演示文稿

第三十九页,共98页。
II 脂肪酸的去饱和:
氧化脱氢途径;光滑型内质网
(1 )单烯脂酸(monoenoic acid)的合成:
人体内有 4, 5, 8, 9去饱和酶,属混合功能氧化酶; 该酶不能在C10与末端甲基之间形成双键
软脂酸 脂酰CoA去饱和酶系
硬脂酸
棕榈油酸 (16, 9 ) 油酸(18, 9 )
合成16个C原子为止。
• 释放:
经7轮cycle合成了棕榈酰-S-ACP
CH3(CH2)14COOH+ACP-SH
CH3(CH2)14CO-S-ACP
第三十二页,共98页。
CH3(CH2)14COSCoA+ACP-SH
软脂酸合成的总反应式:
第三十三页,共98页。
第三十四页,共98页。
丙二酸单酰CoA
第四十页,共98页。
光滑型内质网
NADPH
去饱和酶系:在哺乳动物肝脏和脂肪组织中。
第四十一页,共98页。
(2 )多烯脂酸的形成:植物 (不直接作用于游离脂肪酸)
至今在动物体内尚未发现有 9 以上的去饱和酶 亚油酸(18, 9,12)
必需脂肪酸
(linoleic acid)
-亚麻酸(18, 9,12,15)
包括: 6个酶
酰基载体蛋白(acyl carrier protein, ACP)
•
ACP辅基:4-磷酸泛酰巯基乙胺;摆臂
结合并转运脂酰基
第十七页,共98页。
脂酰基载体蛋白(ACP)的辅基结构
HS -
O-CH2-Ser-ACP
辅基:4-磷酸泛酰巯基乙胺
CoA分子中也有4-磷酸泛酰巯基乙胺
HS
A
4-磷酸泛酰巯基乙胺
第八章合成生物学详解

究项目中,已成功制造出全球首个“合成细胞”,一种 称为丝状支原体的细菌。
第五页,共五十页。
(2) 合成生物学 合成生物学学是生物科学在二十一世纪刚刚出现
的一个分支学科。
目的在于设计和创造新的生物组件和体系,对现有 的生物体系进行重新设计。从基本的生物组件构建复杂 的人工生命体系,对整个生命过程进行重新设计、改造、 构建。
第三页,共五十页。
20世纪提出的概念 用现有的有机化学和生物化学的合成能力设计非天然
的分子,使这些分子在生命体系中发挥功能。通过合成 的方法来理解自然的生命体系,构建创造新的人工生命 体。
第四页,共五十页。
美国基因组学先驱克莱格·凡特,在他位于马里 兰州和加州的实验室,科研人员在其为期15年的研
第八章合成生物学详解演示文稿
第一页,共五十页。
优选第八章合成生物学
第二页,共五十页。
15.1 合成生物学的发展史及概念
(1)合成生物学的发展史
1978年 Skallka在对限制性内切核酸酶的评论中第一次
预言了合成生物学的诞生。
1980年 Hobom引入了合成生物学的的名词来描述基 因重组技术。
DNA合成测序技术的发展和工程学在生物体系的应用, 为合成生物学奠定基础。
红,全身大汗淋漓,大汗后体温降至正常或正常以下。
经过一段间歇期后,又开始重复上述间歇性定 时寒战、高热发作。
第三十页,共五十页。
每年5亿人感染,100万死亡。目前最有效的是青蒿
素,生产周期长、成本昂贵。
中药青篙中提取的有过氧基团的倍半萜内酯药 物。
第三十一页,共五十页。
Keasling利用合成生物学,将大肠杆菌改造成青 蒿酸工厂。将甲羟戊酸合成途径转入大肠杆菌中, 改造获得E. coli 青蒿酸的产量300mg/L。
第五页,共五十页。
(2) 合成生物学 合成生物学学是生物科学在二十一世纪刚刚出现
的一个分支学科。
目的在于设计和创造新的生物组件和体系,对现有 的生物体系进行重新设计。从基本的生物组件构建复杂 的人工生命体系,对整个生命过程进行重新设计、改造、 构建。
第三页,共五十页。
20世纪提出的概念 用现有的有机化学和生物化学的合成能力设计非天然
的分子,使这些分子在生命体系中发挥功能。通过合成 的方法来理解自然的生命体系,构建创造新的人工生命 体。
第四页,共五十页。
美国基因组学先驱克莱格·凡特,在他位于马里 兰州和加州的实验室,科研人员在其为期15年的研
第八章合成生物学详解演示文稿
第一页,共五十页。
优选第八章合成生物学
第二页,共五十页。
15.1 合成生物学的发展史及概念
(1)合成生物学的发展史
1978年 Skallka在对限制性内切核酸酶的评论中第一次
预言了合成生物学的诞生。
1980年 Hobom引入了合成生物学的的名词来描述基 因重组技术。
DNA合成测序技术的发展和工程学在生物体系的应用, 为合成生物学奠定基础。
红,全身大汗淋漓,大汗后体温降至正常或正常以下。
经过一段间歇期后,又开始重复上述间歇性定 时寒战、高热发作。
第三十页,共五十页。
每年5亿人感染,100万死亡。目前最有效的是青蒿
素,生产周期长、成本昂贵。
中药青篙中提取的有过氧基团的倍半萜内酯药 物。
第三十一页,共五十页。
Keasling利用合成生物学,将大肠杆菌改造成青 蒿酸工厂。将甲羟戊酸合成途径转入大肠杆菌中, 改造获得E. coli 青蒿酸的产量300mg/L。
第七讲蛋白质的生物合成ppt文档

缬氨酸、亮氨酸、异亮氨酸
alanine
丙氨酸
Ala
A
arginine
精氨酸
Arg
R
asparagine
天冬酰氨 Asn
N
aspartic acid
天冬氨酸 Asp
D
cystine
半胱氨酸 Cys
C
glutarmine
谷氨酰胺 Gln
Q
glutarmic acid 谷氨酸
Glu
E
glycine
甘氨酸
密码的简并性
一种氨基酸有几组密码子,或者几组密码子代 表一种氨基酸的现象称为密码子的简并性,这种简 并性主要是由于密码子的第三个碱基发生摆动现象 形成的,也就是说密码子的专一性主要由前两个碱 基决定,即使第三个碱基发生突变也能翻译出正确 的氨基酸,这对于保证物种的稳定性有一定意义。 如:GCU,GCC,GCA,GCG都代表丙氨酸。
来源
原核 生物
核糖体 (S)
70
亚基 30
RNA(S) 蛋白质 种类
16
21
50 23, 5 34
图7-2 (a)原核生物mRNA为多顺反子 (b)真核生物mRNA为单顺反子
遗传密码的破译
乔治·伽莫夫(1904~1968 ) (George Gamov) 乌克兰裔美国核物理学家
马歇尔.尼伦贝格(1927-) Marshall Nirenberg 德裔美国生物化学家
奥乔亚(1905-1993) Severo Ochoa
西班牙裔美籍生物化学家
• 1966年: 阐明遗传密码
柯拉那(美国)
Har Gobind Khorana,1922~
霍利(美国)
Robert Holley, 1922-1993
alanine
丙氨酸
Ala
A
arginine
精氨酸
Arg
R
asparagine
天冬酰氨 Asn
N
aspartic acid
天冬氨酸 Asp
D
cystine
半胱氨酸 Cys
C
glutarmine
谷氨酰胺 Gln
Q
glutarmic acid 谷氨酸
Glu
E
glycine
甘氨酸
密码的简并性
一种氨基酸有几组密码子,或者几组密码子代 表一种氨基酸的现象称为密码子的简并性,这种简 并性主要是由于密码子的第三个碱基发生摆动现象 形成的,也就是说密码子的专一性主要由前两个碱 基决定,即使第三个碱基发生突变也能翻译出正确 的氨基酸,这对于保证物种的稳定性有一定意义。 如:GCU,GCC,GCA,GCG都代表丙氨酸。
来源
原核 生物
核糖体 (S)
70
亚基 30
RNA(S) 蛋白质 种类
16
21
50 23, 5 34
图7-2 (a)原核生物mRNA为多顺反子 (b)真核生物mRNA为单顺反子
遗传密码的破译
乔治·伽莫夫(1904~1968 ) (George Gamov) 乌克兰裔美国核物理学家
马歇尔.尼伦贝格(1927-) Marshall Nirenberg 德裔美国生物化学家
奥乔亚(1905-1993) Severo Ochoa
西班牙裔美籍生物化学家
• 1966年: 阐明遗传密码
柯拉那(美国)
Har Gobind Khorana,1922~
霍利(美国)
Robert Holley, 1922-1993
合成生物学简约课件PPT课件

在基因工程的实施过程中由于只转移个 别外源基因,一般较少或不进行细胞网
络分析
改变了“转移一个基因,表达一种蛋白 质”的模式,而通常是转移一组基因, 因而要在更大规模更多层次上涉及到细
胞网络,如代谢网络等
3/28/2020
15
与分子生物学和细胞生物学的关系
• 分子生物学
在分子水平上研究生命现象的科学。通过研究生物大分子 (核酸、蛋白质)的结构、功能和生物合成等方面来阐明各 种生命现象的本质。研究内容包括各种生命过程。比如光 合作用、发育的分子机制、神经活动的机理、癌的发生等
• 2004 年Scientific American 杂志发表了题为
“合成生命”一文, 以图文并茂的形式阐述了合成生物学 的内涵及工程性质
• 2005 年MIT 的Endy发表了“ 工程生物学的基础” 的综 述论文, 明确提出工程中常用的“标准化”、“复杂系 统解耦”、“概念抽象化”做法, 并清楚地将合成生物 学涉及的生物系统分成DNA、零件、装置、系统这样4 个层次
3/28/术2020从专门的工艺转化为一个成熟的工业。
6
• 美国普林斯顿大学电子工程系与分子生物学系的 Weiss等发表了题为“合成生物学: 对于一个刚出现 学科的新的工程作用”的综述。概括了合成生物学 新学科的基本性能以及与其它工程学科相比的独特 性; 讨论了从生物装置、模块、细胞到多细胞系统 各个层次进行设计和建造工程细胞的方法
3/28/2020
22
大肠杆菌砷探测器
• 在世界各地尤其是孟加拉国,饮用水中砷或亚 砷酸盐往往超标,危害数百万人的身体健康。 世界卫生组织(WHO)要求饮用水砷或亚砷酸盐 的浓度不得超过10ppb。现有的砷检测方法难 以达到这一精度。
合成生物学ppt课件

控制生命
目前,研究人员正在试图控制细胞的行为,研 制不同的基因线路———即特别设计的、相互 影响的基因。
波士顿大学生物医学工程师科林斯已研制出一 种“套环开关”,所选择的细胞功能可随意开 关。
加州大学生物学和物理学教授埃罗维茨等人研 究出另外一种线路:
– 当某种特殊蛋白质含量发生变化时,细胞能在发光 状态和非发光状态之间转换,起到有机振荡器的作 用,打开了利用生物分子进行计算的大门。
Venter 实验室发展了合成基因组
Φ X-174 噬菌体基因是单链环状 DNA,是历史上第一个被纯 化的DNA 分子,也是第一个被测序的DNA分子。
Φ X- 174 噬菌体对动植物无害,是合适的合成研究对象。
美国Venter 实验室发展了合成基因组的工作, 该实验室只用 两周就合成了Φ X-174 噬菌体基因 (5,386bp) 。
按照实验计划,最终这个染色体将控制这个细胞并变成一个新的生命 形式。
这种新单细胞生物体被命名为“合成器”,受381个基因控制,包含 56万个碱基对。这些基因是维持细菌生命所必备的,使它能够摄食和 繁殖。由于新的生物体是在现存生物体上搭建,其繁殖和新陈代谢仍 然依赖原来生物体的胞内机制。
从这一角度看,它并非完全意义上的新型生命形式。但这种给特定基 因赋予特定任务的观点已被众多生物学家广泛接受。
What is Synthetic Biology?
synthetic biology
合成生物学(synthetic biology)一词最早出现于1911 年7 月8 日 著名医学刊物《柳叶刀》The Lancet. Reviews and Notices of Books. The Lancet, 1911. 178 (4584):97-99. 发表的一篇书评中。
脂类生物合成演示文稿

第十三页,共27页。
(6) 还原(reduction): 烯酰-ACP还原酶(ER) 产物:丁酰-ACP
进入第二次合成轮回,丁酰-ACP取代第一轮回的
乙酰-ACP(转位(translocation):乙酰CoA:ACP转酰
酶(AT) )
第十四页,共27页。
经过7次循环,螺旋式链增长
(8) 释放(release):脂酰-ACP硫酯酶 释放终产物:软脂酸
内质网的胞液侧。
第二十六页,共27页。
合成原料
甘油-3-P 脂酰-CoA
合成基本过程
1)甘油一酯途径:小肠粘膜细胞(利用消化吸收的甘油
一酯和脂肪酸合成甘油三酯)
2)甘油二酯途径:肝细胞和脂肪细胞
第二十七页,共27页。
第二十二页,共27页。
脂酸碳链的加长
更长碳链的脂酸则是对软脂酸的加工,使其碳链延长。
部位:(肝细胞)内质网或线粒体中
1)内质网脂酸碳链延长酶系
延长过程与软脂酸合成过程相似,但脂酰基连在CoASH上进
行反应.多延长至18个C,最长可至24C。
2)线粒体(延长)酶体系
延长过程与β-氧化的逆行相似,由NADPH+H+供氢,亦 以延长至硬脂酸最多,最长可至24C。
脂类生物合成演示文稿
第一页,共27页。
(优选)脂类生物合成
第二页,共27页。
1.1 原料的转运 软脂酸的合成(C16) 1)合成部位:肝、肾、脑、肺、脂肪线粒体外
2)合成原料:乙酰CoA(主要来自葡萄糖分解)、ATP、
HCO3-、NADPH(主要来自磷酸戊糖途径) 、Mn2+。
原料在线粒体内生成,合成脂肪酸在细胞质,需要将乙 酰CoA运至胞液。
Synthesis
(6) 还原(reduction): 烯酰-ACP还原酶(ER) 产物:丁酰-ACP
进入第二次合成轮回,丁酰-ACP取代第一轮回的
乙酰-ACP(转位(translocation):乙酰CoA:ACP转酰
酶(AT) )
第十四页,共27页。
经过7次循环,螺旋式链增长
(8) 释放(release):脂酰-ACP硫酯酶 释放终产物:软脂酸
内质网的胞液侧。
第二十六页,共27页。
合成原料
甘油-3-P 脂酰-CoA
合成基本过程
1)甘油一酯途径:小肠粘膜细胞(利用消化吸收的甘油
一酯和脂肪酸合成甘油三酯)
2)甘油二酯途径:肝细胞和脂肪细胞
第二十七页,共27页。
第二十二页,共27页。
脂酸碳链的加长
更长碳链的脂酸则是对软脂酸的加工,使其碳链延长。
部位:(肝细胞)内质网或线粒体中
1)内质网脂酸碳链延长酶系
延长过程与软脂酸合成过程相似,但脂酰基连在CoASH上进
行反应.多延长至18个C,最长可至24C。
2)线粒体(延长)酶体系
延长过程与β-氧化的逆行相似,由NADPH+H+供氢,亦 以延长至硬脂酸最多,最长可至24C。
脂类生物合成演示文稿
第一页,共27页。
(优选)脂类生物合成
第二页,共27页。
1.1 原料的转运 软脂酸的合成(C16) 1)合成部位:肝、肾、脑、肺、脂肪线粒体外
2)合成原料:乙酰CoA(主要来自葡萄糖分解)、ATP、
HCO3-、NADPH(主要来自磷酸戊糖途径) 、Mn2+。
原料在线粒体内生成,合成脂肪酸在细胞质,需要将乙 酰CoA运至胞液。
Synthesis
生物合成途径演示文稿

生物合成途径演示文稿
第一页,共20页。
生物合成途径
第二页,共20页。
二、 生物合成途径
第三页,共20页。
薄荷叶为什么牛不吃、虫不咬?
Chemical Defenses
OH
Menthol
menthol broadcast a smell that warns herbivores and insects that the plant is toxic to them
第十三页,共20页。
起始物质为MVA,在ATP作用下, 按如下路线合成。
甲戊二羟酸(MVA) HOOC
O
OH 2ATP
O P2O5H2
焦磷酸二甲烯丙酯
2ADP
ADP ATP
O P2O5H2
焦磷酸异戊烯酯
CO2
HO HOOC
O P2O5H2
甲戊二羟酸-5-焦磷酸
第十四页,共20页。
(三)桂皮酸途径
A
O B
A
O
查耳酮
O
二氢黄酮
第十九页,共20页。
一些萜类生物碱分别来自甲戊二羟酸途 径及莽草酸途径或醋酸—丙二酸途径。
第二十页,共20页。
一级代谢
植物生存 葡萄糖(能量) 必需成分 纤维素(生长)
植物生长
(中药化学成分的来源)
二级代谢
一次代谢
绿色植物以二氧化碳及水为原料,通过 光合作用、固氮反应等生成糖、蛋白质、 脂质、核酸等植物体生命活动必需物质
的过程称为一次代谢过程。 生成的产物
为一次代谢产物。
此外,一次代谢产物还包括乙酰辅酶A,
第四页,共20页。
CORN LEAF BIGHT (VIRUS)
CITRUS CANKER (BACTERIUM)
第一页,共20页。
生物合成途径
第二页,共20页。
二、 生物合成途径
第三页,共20页。
薄荷叶为什么牛不吃、虫不咬?
Chemical Defenses
OH
Menthol
menthol broadcast a smell that warns herbivores and insects that the plant is toxic to them
第十三页,共20页。
起始物质为MVA,在ATP作用下, 按如下路线合成。
甲戊二羟酸(MVA) HOOC
O
OH 2ATP
O P2O5H2
焦磷酸二甲烯丙酯
2ADP
ADP ATP
O P2O5H2
焦磷酸异戊烯酯
CO2
HO HOOC
O P2O5H2
甲戊二羟酸-5-焦磷酸
第十四页,共20页。
(三)桂皮酸途径
A
O B
A
O
查耳酮
O
二氢黄酮
第十九页,共20页。
一些萜类生物碱分别来自甲戊二羟酸途 径及莽草酸途径或醋酸—丙二酸途径。
第二十页,共20页。
一级代谢
植物生存 葡萄糖(能量) 必需成分 纤维素(生长)
植物生长
(中药化学成分的来源)
二级代谢
一次代谢
绿色植物以二氧化碳及水为原料,通过 光合作用、固氮反应等生成糖、蛋白质、 脂质、核酸等植物体生命活动必需物质
的过程称为一次代谢过程。 生成的产物
为一次代谢产物。
此外,一次代谢产物还包括乙酰辅酶A,
第四页,共20页。
CORN LEAF BIGHT (VIRUS)
CITRUS CANKER (BACTERIUM)
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
熟悉:
转录的基本过程;原核生物两类终止子的结构特 点;编码链,模板链;帽子的形成过程;stemloop 结构。
一、转录的概念
转录 (transcription) 是生物体以 DNA为模板合成RNA的过程 。
转录
DNA
RNA
二、复制和转录的异同点
复制
DNA DNA
转录
DNA
RNA
1、复制和转录的相同点
全酶 (holoenzyme)
转录延长阶段
转录起始阶段
RNA聚合酶全酶在转录起始区的结合
三、RNA聚合酶结合到DNA的 启动子上起动转录
• 转录是不连续、分区段进行的。
• 每一转录区段可视为一个转录单位,称为 操纵子(operon)。操纵子包括若干个结构 基因及其上游(upstream)的调控序列。
转录 5′···GCAGUACAUGUC ···3′ mRNA
翻译 N······Ala ·Val ·His ·Val ······C蛋白质
转录的特点
不对称转录(asymmetric transcription)。 它有两方面含义:在DNA分子双链上, 一股链用作模板指引转录,另一股链不 转录;其二是模板链并非总是在同一单 链上。
生物合成详解演示文稿
优选生物合成
本章主要内容OVERVIEW
➢ 转录的概念; ➢ 复制与转录的异同点; ➢ 原核生物RNA聚合酶的组成结构特点; ➢ 真核生物RNA聚合酶的种类及特点; ➢ 原核生物及真核生物启动子结构及特点; ➢ 转录的基本过程; ➢ 真核生物转录后的修饰.
目的要求
掌握:
转录的概念;复制与转录的异同点;原核生物 RNA聚合酶的组成结构;真核生物RNA聚合酶 的种类及转录的产物;启动子的概念,原核生物 及真核生物启动子结构及特点;终止子的概念、 不依赖rho因子的终止子的特点;顺式作用元件, 反式作用因子;真核生物转录后修饰的几种方式; 核酶。
• RNA聚合酶和DNA的特殊序列——启动子 (promoter)结合后,就能启动RNA合成。
(二)RNA聚合酶由多个亚基组成
亚基 α β βˊ σ
分子量
36512 150618 155613 70263
功能
决定哪些基因被转录 催化功能 结合 DNA模板 辨认起始点
核心酶 (core enzyme)
四、转录模板
5′ ···GCAGTACATGTC ···3′ 编码链 3′ ···cgtcatgtacag ···5′ 模板链
转录
5′···GCAGUACAUGUC···3′ mRNA
翻译 N······Ala·Val·His·Val······C 蛋白质
第一节
原核生物转录的模板和酶
Templates & Enzymes in Prokaryotic Transcription
二、RNA合成由RNA聚合酶催化
(一)RNA聚合酶能直接启动RNA链的合成
• 依赖DNA的RNA聚合酶催化合成RNA; • RNA合成的化学机制与DNA依赖的DNA
聚合酶催化DNA合成相似。
( NMP )n + NTP → ( NMP ) n+1 + PPi
RNA
延长的RNA
• DNA聚合酶在启动DNA链延长时需要引物 存在,而RNA聚合酶不需要引物就能直接 启动RNA链的延长。
原料
dNTP
NTP
酶
DNA聚合酶
RNA聚合酶(RNA- pol )
产物
子代双链DNA mRNA,tRNA,rRNA (半保留复制)
配对
A-T,G-C
A-U,T-A,G-C
三、参与转录的物质
原料: NTP (ATP, UTP, GTP, CTP) 模板: DNA 酶: 依赖DNA的RNA聚合酶(RNA polymerase) 其他蛋白质因子
➢ DNA双链解开,使其中的一条链作 为转录的模板。
转录起始
用RNA聚合酶保护法研究转录起始区
RNA聚合酶保护区 结构基因
5
3
3
5
5
3
-50 -40 -30 -20 -10 1 10
3
5
-35 区
TTGACA AA C T G T RNA-pol辨认位点 (recognition site)
开始转录 -10 区
T A T A A T Pu A T A T T A Py (Pribnow box)
调控序列
结构基因
5
RNA-pol
3
3
5
• 调控序列中的启动子是RNA聚合酶结合模 板DNA的部位,也是控制转录的关键部位。 原核生物以RNA聚合酶全酶结合到DNA的 启动子上而起动转录,其中由σ亚基辨认启 动子,其他亚基相互配合。
• 对启动子的研究,常采用一种巧妙 Nhomakorabea方法 即RNA聚合酶保护法。
RNA聚合 酶保护法
一、原核生物转录的模板
DNA双链中按碱基配对规律能指引转录生成RNA的 一股单链,称为模板链(template strand),也称作 有意义链或Watson链。相对的另一股单链是编码 链(coding strand),也称为反义链或Crick链。
5′···GCAGTACATGTC ···3′ 编码链 3′··· c g t c a t g t a c a g ···5′ 模板链
RNA聚合酶全酶在转录起始区的结合:
转录开始的第一个碱基(+1)。原核生物中常 为A或G,而且位置固定。
第二节
原核生物的转录过程
The Process of Transcription in Prokaryote
一、转录起始需要RNA聚合酶全酶
转录起始需解决两个问题:
➢ RNA聚合酶必须准确地结合在转录 模板的起始区域。
结构基因(structural gene)
能转录出mRNA并指导蛋白质合成的DNA片 段。
不对称转录(asymmetric transcription)
结构基因
5
编码链
3
模板链
转录方向
转录方向
模板链
3
编码链
5
➢在DNA分子双链上某一区段,一股链用作 模板指引转录,另一股链不转录 ;
➢模板链并非永远在同一条单链上。
1. 均以DNA为模板 2. 均需要依赖DNA的聚合酶 3. 均以含三个磷酸的核苷酸为原料 4. 均是核苷酸聚合过程,多核苷酸为产物 5. 均遵从碱基配对规律
DNA 3’-ATGCATGC-5’ RNA 5’-UACGUACG-3’
A-U;G-C
2、复制和转录的不同点
复制
转录
模板
两股链均复制 模板链转录(不对称转录)
转录的基本过程;原核生物两类终止子的结构特 点;编码链,模板链;帽子的形成过程;stemloop 结构。
一、转录的概念
转录 (transcription) 是生物体以 DNA为模板合成RNA的过程 。
转录
DNA
RNA
二、复制和转录的异同点
复制
DNA DNA
转录
DNA
RNA
1、复制和转录的相同点
全酶 (holoenzyme)
转录延长阶段
转录起始阶段
RNA聚合酶全酶在转录起始区的结合
三、RNA聚合酶结合到DNA的 启动子上起动转录
• 转录是不连续、分区段进行的。
• 每一转录区段可视为一个转录单位,称为 操纵子(operon)。操纵子包括若干个结构 基因及其上游(upstream)的调控序列。
转录 5′···GCAGUACAUGUC ···3′ mRNA
翻译 N······Ala ·Val ·His ·Val ······C蛋白质
转录的特点
不对称转录(asymmetric transcription)。 它有两方面含义:在DNA分子双链上, 一股链用作模板指引转录,另一股链不 转录;其二是模板链并非总是在同一单 链上。
生物合成详解演示文稿
优选生物合成
本章主要内容OVERVIEW
➢ 转录的概念; ➢ 复制与转录的异同点; ➢ 原核生物RNA聚合酶的组成结构特点; ➢ 真核生物RNA聚合酶的种类及特点; ➢ 原核生物及真核生物启动子结构及特点; ➢ 转录的基本过程; ➢ 真核生物转录后的修饰.
目的要求
掌握:
转录的概念;复制与转录的异同点;原核生物 RNA聚合酶的组成结构;真核生物RNA聚合酶 的种类及转录的产物;启动子的概念,原核生物 及真核生物启动子结构及特点;终止子的概念、 不依赖rho因子的终止子的特点;顺式作用元件, 反式作用因子;真核生物转录后修饰的几种方式; 核酶。
• RNA聚合酶和DNA的特殊序列——启动子 (promoter)结合后,就能启动RNA合成。
(二)RNA聚合酶由多个亚基组成
亚基 α β βˊ σ
分子量
36512 150618 155613 70263
功能
决定哪些基因被转录 催化功能 结合 DNA模板 辨认起始点
核心酶 (core enzyme)
四、转录模板
5′ ···GCAGTACATGTC ···3′ 编码链 3′ ···cgtcatgtacag ···5′ 模板链
转录
5′···GCAGUACAUGUC···3′ mRNA
翻译 N······Ala·Val·His·Val······C 蛋白质
第一节
原核生物转录的模板和酶
Templates & Enzymes in Prokaryotic Transcription
二、RNA合成由RNA聚合酶催化
(一)RNA聚合酶能直接启动RNA链的合成
• 依赖DNA的RNA聚合酶催化合成RNA; • RNA合成的化学机制与DNA依赖的DNA
聚合酶催化DNA合成相似。
( NMP )n + NTP → ( NMP ) n+1 + PPi
RNA
延长的RNA
• DNA聚合酶在启动DNA链延长时需要引物 存在,而RNA聚合酶不需要引物就能直接 启动RNA链的延长。
原料
dNTP
NTP
酶
DNA聚合酶
RNA聚合酶(RNA- pol )
产物
子代双链DNA mRNA,tRNA,rRNA (半保留复制)
配对
A-T,G-C
A-U,T-A,G-C
三、参与转录的物质
原料: NTP (ATP, UTP, GTP, CTP) 模板: DNA 酶: 依赖DNA的RNA聚合酶(RNA polymerase) 其他蛋白质因子
➢ DNA双链解开,使其中的一条链作 为转录的模板。
转录起始
用RNA聚合酶保护法研究转录起始区
RNA聚合酶保护区 结构基因
5
3
3
5
5
3
-50 -40 -30 -20 -10 1 10
3
5
-35 区
TTGACA AA C T G T RNA-pol辨认位点 (recognition site)
开始转录 -10 区
T A T A A T Pu A T A T T A Py (Pribnow box)
调控序列
结构基因
5
RNA-pol
3
3
5
• 调控序列中的启动子是RNA聚合酶结合模 板DNA的部位,也是控制转录的关键部位。 原核生物以RNA聚合酶全酶结合到DNA的 启动子上而起动转录,其中由σ亚基辨认启 动子,其他亚基相互配合。
• 对启动子的研究,常采用一种巧妙 Nhomakorabea方法 即RNA聚合酶保护法。
RNA聚合 酶保护法
一、原核生物转录的模板
DNA双链中按碱基配对规律能指引转录生成RNA的 一股单链,称为模板链(template strand),也称作 有意义链或Watson链。相对的另一股单链是编码 链(coding strand),也称为反义链或Crick链。
5′···GCAGTACATGTC ···3′ 编码链 3′··· c g t c a t g t a c a g ···5′ 模板链
RNA聚合酶全酶在转录起始区的结合:
转录开始的第一个碱基(+1)。原核生物中常 为A或G,而且位置固定。
第二节
原核生物的转录过程
The Process of Transcription in Prokaryote
一、转录起始需要RNA聚合酶全酶
转录起始需解决两个问题:
➢ RNA聚合酶必须准确地结合在转录 模板的起始区域。
结构基因(structural gene)
能转录出mRNA并指导蛋白质合成的DNA片 段。
不对称转录(asymmetric transcription)
结构基因
5
编码链
3
模板链
转录方向
转录方向
模板链
3
编码链
5
➢在DNA分子双链上某一区段,一股链用作 模板指引转录,另一股链不转录 ;
➢模板链并非永远在同一条单链上。
1. 均以DNA为模板 2. 均需要依赖DNA的聚合酶 3. 均以含三个磷酸的核苷酸为原料 4. 均是核苷酸聚合过程,多核苷酸为产物 5. 均遵从碱基配对规律
DNA 3’-ATGCATGC-5’ RNA 5’-UACGUACG-3’
A-U;G-C
2、复制和转录的不同点
复制
转录
模板
两股链均复制 模板链转录(不对称转录)