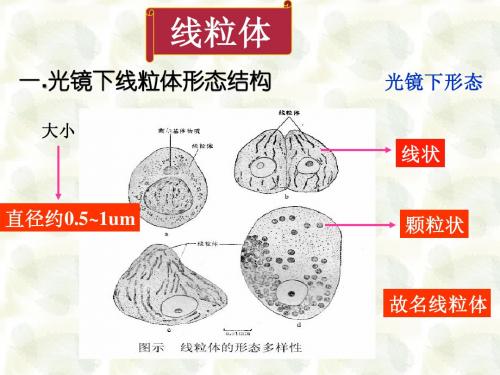
线粒体的形态结构.
细胞生物学 线粒体

Arg
H+通道I
γ和ε的转动引起β构象变化,催化ATP生成
结合变化机制:
β三种状态:
O 空置状态
ADP+Pi ATP
T 紧密结合态
L 松散结合态
转子旋转的试验学证据:
• 在嗜热菌的 F1ATPase β的N端 添加10个His,固定 在Ni-NTA膜上; • 将γN端的一个Ser换 为Cys,以连上肌动 蛋白丝(荧光标记); • 加ATP后可观察到 肌动蛋白丝旋转;
基粒(elementary particle)
• 内膜和嵴的基质面上带柄的颗 粒; • 由头部(F1)和基部(F0)构 成。 (F1-F0 ATP酶)
长 轴 垂 直 板 层 状 嵴
管状嵴
纵行排列板层状嵴
心肌细胞排列紧密的板层状嵴
3 膜间腔
• 外膜与内膜间的空腔,由于外膜通透性很强,
而内膜的通透性又很低,所以膜间隙中的化学
3)呼吸链的电子传递及其抑制剂和作用部位
鱼藤酮 ×
氰化物 叠氮钠 CO
×
×
抗霉素A
氧化还原电势
琥珀酸 脱氢酶
通过电子传递链实现了跨膜的H+浓度差
2 ATP合成酶 (ATP synthase)
F1:亲水性头部,易分离,催 线粒体基质 化ATP ,含3α、3β、γ、δ和ε; • 水解 的多亚基蛋白复合物, β:催化 ATP合成 水解; 由水溶性 F1/ 头部和疏水 ε:抑制 水解,堵塞H+通道; 性ATP F0基部所组成,也叫 F1F0 ATPase。 F0:疏水性基部,传送H+到F1, F0 含1a, 2b, 10-14c。 a: H+通道; “转子” :γ,ε
◆复合物Ⅰ:NADH-CoQ还原酶复合物(电子传递 体和H+ 移位体) 组成:含42个亚基,至少6Fe-S中心和1黄素蛋白。 作用:氧化NADH,将2e-CoQ;泵出4 H+ ◆复合物Ⅱ:琥珀酸脱氢酶复合物(电子传递体而 非H+移位体) 组成:含FAD辅基,2Fe-S中心,1Cytb 作用:氧化琥珀酸,2e-FADFe-SCoQ
线粒体-细胞生物学

线粒体形态结构:椭圆形少数圆形宽0.8~1μm,长1~2μm(特殊7~40μm)两层单位膜外膜、内膜、外室、内室组成内膜向线粒体腔形成褶叠,称Mit嵴(嵴间平行,与纵轴垂直)嵴增加内膜面积(外膜的5倍,细胞膜结构总面积的1/3)基粒(F1颗粒、F1因子、内膜亚单位)外膜、内膜、外室、内室组成线粒体化学组成蛋白质、脂、水(脂:磷脂等膜脂;水:代谢产生)蛋白质最重要成份可溶性:基质酶、膜外周蛋白不溶性:内外膜整合蛋白(受体、酶等)蛋白质在线粒体各部分分布不同外膜两类:►(1)氧化还原酶:单胺氧化酶、NADH-细胞色素C还原酶等(单胺氧化酶是外膜标志酶)►(2)脂类代谢酶:酰基辅酶A合成酶、脂肪酸延伸酶等此外:孔蛋白,2~3nm通道,外膜具有通透性,小分子物质自由进出外室中酶种类少,主要为激酶,如腺苷酸激酶(标志酶)、二磷酸激酶等内膜上酶种类多、结构复杂、许多酶以复合物的形式出现内膜上的酶,四类:(1)移位酶:特异性载体,运输磷酸、Ca2+、核苷酸、α-酮戊二酸、谷氨酸、鸟氨酸等代谢产物(2)生物合成酶类:参与Mit DNA、RNA、蛋白质合成、脂肪酸、血红素合成(3)呼吸链酶:构成内膜的主要成份a.复合物I:NADH脱氢酶b.复合物II: 琥珀酸脱氢酶c.复合物III:CoQ-细胞色素C还原酶d.复合物IV:细胞色素氧化酶(4)能量转换酶——基粒(ATP酶复合体)♦基粒:ATP合成酶(F1-F0偶联因子,F1-F0 ATP酶)三部分:头部:球状,直径9nm,容易分离,水溶性,称F1因子,由α3β3εδγ 9亚基组成►分离的F1因子能催化:ADP+Pi → ATP也能催化:ATP → ADP+Pi►但Mit内,合成反应,存在一种热稳定蛋白(MW1万),可与F1结合,抑制ATP水解,促进合成►称F1抑制蛋白柄部:►由一种蛋白质组成►高4.5nm,容易分离►作用:使F1因子对寡霉素敏感►F1因子合成ATP的活性能被寡霉素抑制,但寡霉素不直接作用于F1因子,通过柄部蛋白传递►称寡霉素敏感性转授蛋白(oligomycin-sensitivity confering protein,OSCP)膜部:►嵌合在膜内疏水蛋白复合体,称F0因子►至少由3~4条肽链组成►其中一种与寡霉素结合,通过OSCP对F1因子起作用►与质子H+传递有关►当去除F1 和OSCP,内膜对H+的通透性↑,结合上去,可堵住H+泄漏,推测有H+通道线粒体ATP的合成►ATP合成是线粒体最重要功能►ATP来源于糖、氨基酸、脂肪酸的氧化►三类物质存在共同的氧化途径:三羧酸循环ATP合成几种假说:►化学偶联假说►构象偶联假说►化学渗透假说(1978年诺贝尔奖)又称电化学偶联假说基本观点:呼吸链在电子传递中,起着质子泵的作用从NADH脱氢酶开始,每传递一对电子(2e),将线粒体基质的3对质子(6H+)泵到外室(从琥珀酸脱氢酶开始?)化学渗透假说►随着电子传递不断进行,内膜内外形成[H+]浓度差和电位差►电化学梯度(势能)►外室[H+]有返回基质的趋势►每一对质子,驱动合成一个ATP实验证明:微电极,内膜两侧存在电位和pH梯度缬氨酶素,内膜通透性↑,[H+]梯度消失,电子传递进行,但ATP 合成停止►ATP合成酶工作机制►分子“马达”►旋转催化机制►γ亚基像车轮的“轴”,在质子流推动下,转动►带动与“轴”相连的3个β亚基转动(逆时针)►导致β亚基构象交替变化(三种构象:L 结合ADP+Pi;T 合成ATP;O 释放ATP,并周而复始)►1997年,诺贝尔化学奖线粒体的增殖和起源进化(1)增殖分割或分裂产生(2)起源进化内共生假说细菌被原始真核细胞吞噬,演变成线粒体(叶绿体蓝藻)非共生假说共生假说证据:Mit内膜与细菌质膜相似,表现在:►都有呼吸链酶►细菌质膜可以向细胞质内延伸出类似于线粒体嵴的结构(中体)Mit外膜与真核细胞质膜相似(内吞时质膜包围)►Mit DNA与细菌DNA相似:双链环状,复制方式和过程,分子量,裸露►核糖体相似:沉降系数70S左右,小亚基16S rRNA核苷酸序列相似,蛋白质合成过程相似,各种抗生素抑制抗生素抑制细菌生长机理:抑制70S核糖体(不抑制80S核糖体)。
_线粒体的形态,数目和结构资料

总之:线粒体的形态、大小、 数目和分布在不同形态和类型 的细胞中可朔性较大。
一般代谢旺盛细胞中线粒体多,反之则少;
鼠动脉平滑肌细胞 细胞核染成蓝色,绿 色示线粒体,红色示 肌动蛋白纤维 线粒体围绕着精子 尾部鞭毛的中轴
●单细胞鞭毛藻中1个线粒体 ●巨大变形虫中约50万个线粒体 ●哺乳动物肝细胞中约2000个线粒体,肾细胞中约300个,
精子中约25个
分布:因细胞形态和类型的不同而存在差异。通常分布 于细胞生理功能旺盛的区域和需要能量较多的部位。
●精细胞中,沿鞭毛紧密排列;肌细胞中,包装在邻近肌原纤维中间
细胞内线粒体分布可因细胞的生理状态改变产生移位现象
位于外膜内侧,由一层单位膜构成,平均厚4.5nm。含100种以 上的多肽,蛋白质和脂类的比例约4:1。心磷脂含量高(达 20%)、缺乏胆固醇,类似于细菌。其通透性很差,但有高度的 选择通透性,借助载体蛋白控制内外物质的交换。内膜向内形成 嵴,嵴增大了线粒体内膜的表面积,嵴的内表面上布满了颗粒— —基粒。基粒由头部(F1)和基部(F0)构成 。
板层状嵴线粒体
管状嵴线粒体
基粒:内膜和嵴膜基质面上许多带柄的小颗粒。每个
线粒体约104~105个。与膜面垂直而规律排列。 基粒是将呼吸链电子传递中释放的能量用于使ADP 磷酸化生成ATP的重要部位(ATP合酶复合体)。 头部: 合成ATP 柄部: 调节质子通道 基片:质子的通道
可溶性的ATP酶(F1)既 是合成酶又是水解酶
光 镜 下 线 粒 体 的 结 构
电 镜 下 的 形 态
短棒状 或小球状
线粒体是由双层单位膜包围而形式的囊状小体
线粒体的结构ppt课件

三、 线 粒 体 的 功 能
线粒体主要功能是:进行氧化磷酸化,合 成ATP,为细胞生命活动提供直接能量。
细胞氧化:指依靠酶的催化,氧将细胞内
各种供能物质氧化而释放能量的过程。由于
细胞氧化过程中,要 所以又称为细胞呼吸
消耗
O2
数目多变性: 不同细胞中数目不同,约1--50万个不等。 一般代谢旺盛细胞中线粒体多,反之则少;
鼠动脉平滑肌细胞 细胞核染成蓝色,绿 色示线粒体,红色示 肌动蛋白纤维
线粒体围绕着精子 尾部鞭毛的中轴
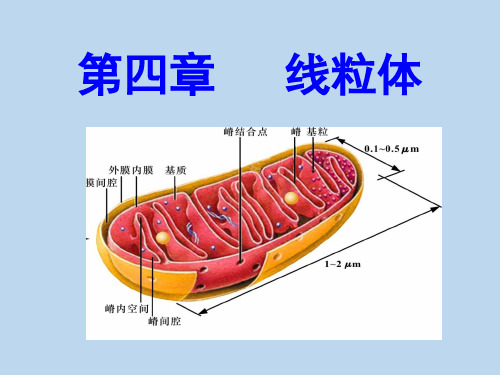
线粒体超微结构:
外膜 内膜 膜间腔(外腔) 基质(内腔)
嵴的形态多样
羽冠型 网膜型 绒毛型 平行型 同心园型
线粒体异常导致的疾病: 如:KSS 综合症:眼肌麻痹、小脑、心脏 异常 MELAS 综合症:中风样发作、癫痫、痴呆 LEBER 遗传性视神经精病:视神精坏 线粒体心肌病:心力衰竭、全身肌无力伴
水肿等、周围视力存在 帕金森病:震颤性麻痹,痴呆等。
利用呼吸链组分进行疾病治疗: Cytc 治疗CO中毒、缺氧窒息、心肺功能障碍 ATP 治疗体质虚弱 CoQ 治疗牙周病、高血压、肌肉萎缩症 NAD+ 治疗进行性肌萎缩、某些肝病. 毒物和药物可作用于呼吸链引起中毒 一氧化碳中毒(缺氧、呼吸链受阻)
Mi内膜与基粒
能量生成
1葡萄糖 酵解 2ATP+2丙酮酸
2丙酮酸
氧化 磷酸化
34ATP 2ATP
38ATP
四、线粒体的半自主性
线粒体有独立的遗传系统
1.mtDNA是环状,裸露,信息量较 小,有独立的编码系统,和细菌DNA 相似。 2.mtDNA可进行自我复制,转录自 己的mRNA、tRNA、rRNA 3.有自己的核糖体,能独立合成线 粒体蛋白质(电子传递链酶复合体 中的亚基:细胞色素C氧化酶、ATP 酶复合体F0的亚基等) 4. mtDNA所用遗传密码和“通用” 的遗传密码不完全相同。
线粒体

04.06.2020
精选
33
三羧酸循环
线粒体基质中乙酰辅酶A与草酰乙酸缩合形成柠檬酸
柠檬酸经一系列反应氧化脱羧,生成草酰乙酸
草酰乙酸与另一分子的乙酰辅酶A结合重新形成柠檬酸
净生成2分子GTP,8对H原子,6对由NAD+携带,2对由
04.0F6.A20D20 携带
精选
34
三羧酸循环
苹果酸
乙酰辅酶A 草酰乙酸
内 膜(inner membrane)
膜间腔(intermembrane space)
基质(matrix)
04.06.2020
精选
12
04.06.2020
线粒体超微结构: 外膜 内膜 膜间腔(外腔) 基质(内腔)
精选
13
04.06.2020
基质(内腔)
嵴
膜间腔(外腔)外) 内腔(与基质相通) (内腔) 内膜
精选
51
ATP合成酶的分子结构及作用机制
美国生物化学家Boyer(1989)提出了结合变构机制(binding-change mechanism)来解释ATP合成酶如何利用跨膜的质子梯度形成ATP
F1和F0通过“转子”和“定子”连接起来,在合成ATP的过程中,“转子” 在H+流推动下旋转,调节3个β亚基的构象变化。
04.06.2020
精选
43
化学渗透假说
1961年Peter D. Mitchell 提出
线粒体内膜对H+是不通透的,内膜中的呼吸链起着 质子泵的作用,在内膜两侧形成电化学质子梯度, 当H+沿ATP酶复合体到基质时,ATP合成,实现氧 化磷酸化偶联
特点 强调膜结构的完整性
04.06.2020
线粒体-药学细胞学
2.
自主性的限制
(1)mtDNA 信息量少,只能合成5%的内膜蛋白。 mtDNA编码: 2种rRNA(构成线粒体核糖体) 22种tRNA(线粒体转运 RNA) 13种多肽(只形成呼吸酶复合体的6个亚单位) (2)其蛋白合成系统中的DNA聚合酶,RNA聚合酶, 核糖体蛋白质、氨基酸活化酶等仍由核基因编码。
细胞呼吸(细胞氧化)过程
糖酵解
乙酰辅酶A的形成
三羧酸 循环 电子传递和 氧化磷酸化
图示 细胞呼吸的四个主要步骤
葡萄糖分解成丙酮酸 葡萄糖磷酸化成6-磷酸葡萄糖
6-磷酸葡萄糖转变成6-磷酸果糖 6-磷酸果糖转变成1,6-二磷酸果糖 1,6-二磷酸果糖裂解成磷酸二羟丙酮和3-磷酸甘油醛 3-磷酸甘油醛氧化成1,3-二磷酸甘油酸 1,3-二磷酸甘油酸转变成3-磷酸甘油酸 3-磷酸甘油酸转变成2-磷酸甘油酸 2-磷酸甘油酸转变成磷酸烯醇式丙酮酸 磷酸烯醇式丙酮酸转变成丙酮酸
电子传递和氧化磷酸化 1分子NADH —————————— 3ATP 电子传递和氧化磷酸化 1分子FADH2 —————————— 2ATP 所以 10NADH+2FADH2 34ATP
底物水平磷酸化 由高能底物水解放能,直接将高能磷酸链从底物转移 到ADP上,使 ADP磷酸化生成 ATP的作用,称为底物水 平磷酸化。 在糖酵解时,由底物水平磷酸化,产生2ATP(细胞质中 产生); 在TAC时,由底物水平磷酸化,也产生2ATP 。
细胞氧化
1分子葡萄糖 38分子ATP 利用率40.4%
(3)电子传递和氧化磷酸化偶联机制
1961年Mitchell的化学渗透学说
电子传递过程中释放的能量形成质子泵,三次把H+ 从膜内泵出膜外,进入线粒体的膜间腔及嵴间腔 造成内膜两侧的质子浓度差, H+顺电化学差从质子 通道渗透到线粒体的基质中,所释放的自由能驱动 F0F1酶复合体合成ATP.
细胞生物学第七章 线粒体ppt课件

■ 两套遗传体系的协同性
通过离体实验发现两套 遗传体系的遗传机制不 同。 如放线菌酮是细胞质蛋 白质合成抑制剂,但是 对细胞器蛋白质的翻译 却没有作用。另外,一 些抗生素,如氯霉素、 四环素、红霉素等能够 抑制线粒体蛋白质的合 成,但对细胞质蛋白质 合成没有多大影响。 通过对转录的抑制研究, 发现线粒体基因转录的 RNA聚合酶也是特异 的(图)。
线粒体蛋白转运
图 线粒体蛋白转运的部位
分子伴侣(molecular chaperon)
概念:一类在序列上没有相关性但有共同功能的蛋白质,它 们在细胞内帮助其他含多肽的结构完成正确的组装,而且在 组装完毕后与之分离,不构成这些蛋白质结构执行功能时的 组份。 种类:伴侣素家族(chaperonin, Cpn)、热休克蛋白 家族 ( Hsp family )、 核质素、T 受体结合蛋白 (TRAP) 等 特征:1、分子伴侣对靶蛋白没有高度专一性,同一分子伴 侣可以促进多种氨基酸序列完全不同的多肽链折叠成为空间 结构、性质和功能都不相关的蛋白质。 2、它的催化效率很低。行使功能需要水解ATP,以改 变其构象,释放底物,进行再循环。 3、它和肽链折叠的关系,是阻止错误折叠,而不是促 进正确折叠。 4. 多能性(胁迫保护防止交联聚沉,转运,调节转录 和复制,组装细胞骨架) 5. 进化保守性
细胞生物学第七 章 线粒体
第一节、 线粒体的生物学特征
线粒体是能够在光学显微镜进行 观察的显微结构。 ● 1890年,德国生物学家 Altmann第一个发现线粒体。 ● 1897年对线粒体进行命名。 ● 1900年,Leonor Michaelis用 染料Janus green对肝细胞进行 染色,发现细胞消耗氧之后,线 粒体的颜色逐渐消失了,从而提 示线粒体具有氧化还原反应的作 用。
第六章 线粒体

◆ 辅酶Q(CoQ)、黄素单核苷酸(FMN)、 黄素腺嘌呤二核苷酸(FAD)、烟酰胺腺 嘌呤二核苷酸(NAD)等。它们作为辅酶 (或辅基)参与电子传递的氧化还原过程。
◆ 基质中含有催化三羧酸循环、脂肪酸β-氧 化、氨基酸氧化、蛋白质合成等有关的上 百种酶和其他成分, 如环状DNA、RNA、 核糖体及较大的致密颗粒,这些颗粒是含 磷酸钙的沉积物,其作用是储存钙离子, 也可结合镁离子。基质中还有许多可溶性 代谢中间物。
化学渗透假说有两个特点:
A. 强调线粒体膜结构的完整性
如果膜不完整,H+ 便能自由通过膜,则无法在内 膜两侧形成质子动力势,那么氧化磷酸化就会解 偶联。一些解偶联剂的作用就在于改变膜对H+ 的 通透性,从而使电子传递所释放的能量不能转换 合成ATP。
B. 定向化学反应
ATP水解时,H+从线粒体内膜基质侧抽提到膜间 隙,产生电化学质子梯度。ATP合成的反应也是 定向的,在电化学质子梯度推动下,H+ 由膜间隙 通过内膜上的ATP合成酶进入基质,其能量促使 ADP和Pi合成ATP。
◆ 复合物Ⅳ:细胞色素C氧化酶
组成: 二聚体,每一单体含13个亚基,含cyt a, a3 ,Cu, Fe。既是电子传递体又是质子移位 体。 作用: 催化电子从cyt c分子O2 形成水,2 H+泵 出, 2 H+ 参与形成水。
在电子传递过程中,有几点需要说明
◆ 四种类型电子载体:黄素蛋白、细胞色素(含血红 素辅基)、 Fe-S 中心和辅酶 Q。前三种与蛋白质 结合,辅酶Q为脂溶性醌。 ◆ 电子传递起始于NADH脱氢酶催化NADH氧化,形 成高能电子(能量转化),终止于O2形成水。 ◆ 电子传递方向按氧化还原电势递增的方向传递 (NAD+/NAD最低,H2O/O2最高)。
线粒体

-
踏 车
微管蛋白 微管蛋白
聚合
异二聚体
首尾相连
原纤维
(13)
微管
1 13 12 11
2
3 4 5 6
10
7 9 8
F1
F1 由5个不同的亚单位组成——3 3 1 1 1。 交替环状排列,形 成球形头部; 形成柄部, 贯穿 于头部并形成柄的一部分。
F0 由3个不同的亚单位组成—— 1a 1b 12c。
F0
c 亚单位形成膜上可移动的环; a 亚单位可作为质子的通道; b 亚单位嵌入脂质双层,并延伸到F1, 形成柄的一部分。
线粒体 (Mitochondria, Mi)
是细胞内一种重要的细胞器,是细胞内的能量转 换系统,通过氧化磷酸化产生的ATP 为细胞生命活动 提供能量。生物体80%的ATP由Mi产生,故Mi被称为 细胞的“动力工厂”。
一、Mi的形态结构
(一) 形态、大小、数目和分布
形态:多种多样,常见为杆状或颗粒状。
Treadmilling
-
+
影响微丝聚合与解聚的特异性药物与离子:
细胞 松弛素:特异性的破坏微丝组装。
鬼笔环肽:稳定微丝、促进微丝聚合。
在含:ATP和Ca2+、高浓度的单价离子(Na+、K+ 等)溶液中G-actin F-actin
肌钙蛋白
原肌球蛋白
单体
四.微丝的功能
线粒体
A 羽冠型
B 网膜型
C 绒毛型
D 平行型
E 同心圆型
扁层状嵴线粒体
扁层状嵴线粒体
管状嵴线粒体
2.基粒(ATP合酶/ 基粒是氧化磷酸化的结构部位, F0F1ATP酶) 其化学本质是F0F1ATP合成酶
ADP+Pi 头部(ATP酶复合体) ATP
柄部:调控质子通道
基片:为疏水蛋白, 它是质子的通道。
第四章
线粒体
第一节 线粒体的形态、 数量和结构
一、线粒体的形态结构
形状:线粒体一般呈粒状或杆状,环形,哑铃形、 线状、分杈状或其它形状。
线粒体的形态和细胞的种类和细 胞所处的生理状态不同有关。
成纤维细胞线条状线粒体 家兔肝脏细胞颗粒状线粒体
一、线粒体的形态结构
大小:一般直径0.5~1μm,长1.5~3.0μm,在人的 成纤维细胞中可长达40μm,称巨线粒体。
非折叠状态 NAC:与少数前体蛋白相互作用,增加蛋白转运的
准确性。
hsc70:和绝大多数的前体蛋白结合,使前体蛋白
打开折叠,防止已松弛的前体蛋白聚集。
二、转运过程
1、前体蛋白在线粒体外保持非折叠 状态、与受体结合。
2、分子运动产生的动力协助多肽链
穿越线粒体膜;
3、多肽链在线粒体基质内重新折叠。数量:数百至源自千新陈代谢旺盛的细胞,线粒体多
肿瘤细胞呼吸能力弱,线粒体较正常细胞少
分布:均匀分布在细胞质中
分布在需要供能的部位
线粒体的电镜照片
二、线粒体的超微结构
线粒体(mitochondrion)是由两层单位膜套叠而成 的封闭的囊状结构。
二、线粒体的超微结构
线粒体(mitochondrion)是由两层单位膜套叠而成 的封闭的囊状结构。
大学医学细胞生物学线粒体

殖
形成两个新线粒体。
出芽分裂 :先从线粒体上长出小芽,然后小芽与母线粒体分
离,经过不断长大,形成新的线粒体。
线粒体的增殖
间壁分离
收缩分离
出芽分裂
线粒体的起源
内共生假说:线粒体是由共生于原始真核细胞内的细菌演变而来。
依据:
线
1.线粒体DNA呈环状、裸露与细菌相似。
粒 体
2.线粒体的核糖体为70S与细菌相同,而真核细胞为80S。
关于NADH氧化呼吸链
• NAD+ 为氧化型烟酰胺腺嘌呤二核甘酸(即 氧化型辅酶I)
• NADH为还原型烟酰胺腺嘌呤二核甘酸(即 还原型辅酶I)
• 生物氧化过程中大多数脱氢酶都以NAD+ 为 辅酶,也就是说,底物脱下的氢首先被NAD+ 接受,然后再向下传递。
• 复合体I:即NADH-泛醌还原酶(NADH脱氢酶复 合体),可将电子传递给泛醌。该复合物含有以 FMN为辅基的黄素蛋白和以Fe-S为辅基的铁硫蛋 白。
2 线粒体内膜本身具有离子不通透性,能隔绝包括H+、OH-在内 的各种正负离子,正是由于线粒体内膜是质子屏障,产生跨膜 的电化学梯度,外室中高浓度的H质子有返回内室的趋势。
3 当H质子从外室通过基粒F0上的质子通道进入内室时,驱动F1 因子旋转, 利用这种势能使ADP磷酸化合成ATP。
一、线 粒 体 的DNA ( mtDNA )
基粒:内膜和嵴膜内表面上附着带柄的颗粒,亦称为“ATP
合成酶复合体”。它是氧化磷酸化最终产生ATP的部位。
(四)基质
内膜和嵴围成的腔隙,
嵴间腔 (内室)
腔内充满均质的胶状
物质——基质。
膜间隙 (外室)
嵴 内膜 外膜
线粒体

化学渗透假说详细示意图
胞液侧 H+ + + + + + + Q
H+ Cyt c H+
+
+ +
F
+
NADH+H+
Ⅰ
Ⅱ
NAD+ 琥珀酸
延胡索酸
Ⅲ
Ⅳ
0
- - 1/2O2+2H+
- H 2O F1
-
基质侧
ADP+Pi H+ ATP
质子动力势 ( △P)
动画1
动画2
3个ATP合成部位 个 合成部位
1分子葡萄糖产生的能量
在线粒体基质中进行,二十几步酶促化学反应 。
结果: 2×CH2CO~SCOA→2×2CO2 →2×3NADH+H+ →2×1FADH2 → 2×1ATP
分子CO2,能量已转移到 NADH、FADH2、 ATP中
即 G →10 → →2 FADH2 →4ATP →6 CO2 下面的问题是 NADH、 FADH2中的能 NADH、
(一)氧化磷酸化的分子结构基础
1968年,E Racker等 人用超声波将线粒体 破碎,线粒体内膜可 自然卷曲成颗粒朝外 的小膜泡,——亚线 亚线 粒体小泡or亚线粒体 粒体小泡 亚线粒体 颗粒。具有正常的电 颗粒 电 子传递和磷酸化 磷酸化的功 子传递 磷酸化 能。
此实验说明:
参与氢和电子传递的各种氧化还原酶即呼 呼 相当于“ 吸链位于线粒体内膜中。相当于“放能装 相当于 吸链 氧化的结构基础) 置”。(氧化的结构基础 氧化的结构基础 而颗粒 颗粒是氧化磷酸化的耦联因子,位于内 颗粒 膜的基质侧,它是ATP合酶的组分之一, 相当于“换能装置”。(磷酸化的结构基 相当于“换能装置”。(磷酸化的结构基 础)
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
葡萄糖
细
2ATP
胞 氧
丙酮酸
化
CO2
第三节 线粒体的功能 ⑹
氧化磷酸化(oxidative phosphorylation) ——在电子传递过程中,氧化所释放出来的能 量被磷酸化转换成ATP(ADP+Pi+能量 →ATP),这种伴随电子传递链的氧化过程所 进行的能量转换和ATP的生成就是氧化磷酸化。 该过程产生34ATP。 线粒体内形成ATP,并贮存能量。1分子 葡萄糖彻底氧化生成CO2和H2O,净产生 38 ATP。
第一节 线粒体的形态结构 ⑷
⑵ 内膜(inner membrane) ——位于外膜的内侧,一层单位膜,厚
5~6nm,蛋白质与脂类的比例为3.8∶1。 ⑶ 嵴(cristae)
——内膜向内折叠形成,有管状,板层状两 种类型。
第一节 线粒体的形态结构 ⑸
⑷ 基粒(elementary particle) ——位于内膜和嵴的基质面的带柄的圆形颗粒,
㈡ 大小:一般直径在0.5~1.0μm,长3μm, 最长的8~10μm。与细胞种类、生理 状态及环境条件有关。
第一节 线粒体的形态结构 ⑵
㈢ 数目:正常细胞一般有1000~2000个 线粒体。与细胞种类和生理状态 有关。
㈣ 分布:与细胞种类和生理状态有关。线 粒体的分布有一定的规律,通常 分布在功能旺盛,需能较多的部 位。
二. 酶的分布 线粒体中约有120种酶(参见表8-1) 线粒体各部分的标志酶 外膜——单胺氧化酶 膜间腔——腺苷酸激酶 内膜——琥珀酸脱氢酶 基质——苹果酸脱氢酶
第三节 线粒体的功能 ⑴
主要功能——细胞氧化和能量转换 细胞氧化(cellular oxidat (糖、脂肪、氨基酸等)氧化并释放能量的过 程。也称细胞呼吸(cellular respiration)。 能量转换 ——在线粒体内膜上(电子传递过程中),将 氧化反应释放的能量转化成细胞可利用的ATP的 过程。
F1
F0
• F1 由5种不同的亚单位组成——3 3 1 1 1。 交替环状排列,形 成球形头部; 形成柄部, 贯穿 于头部并形成柄的一部分。
• F0 由3个不同的亚单位组成—— 1a 2b 12c。
c 亚单位形成膜上可移动的环; a 亚单位可作为质子的通道; b 亚单位嵌入脂质双层,并延伸到F1,
⑺ 嵴内腔(intracristal space) ——由嵴膜包围,与膜间腔相通。
第一节 线粒体的形态结构 ⑺
⑻ 基质(matrix) ——内腔中所充满的较致密的物质。
⑼ 基质颗粒(matrical granule) ——位于基质中的一种较大的颗粒,能调节 线粒体内离子环境。
⑽ 核糖体 ——存在于内腔中。
第三节 线粒体的功能 ⑵
细胞氧化过程包括—— 糖酵解 乙酰CoA生成 三羧酸循环 电子传递和氧化磷酸化
氧化偶联磷酸化产生ATP
Glucose
第三节 线粒体的功能 ⑶
㈠ 糖酵解:在细胞质基质中进行,脱掉2H进入呼吸 链
酵解酶系
1分子葡萄糖
2分子丙酮酸+2ATP
㈡ 乙酰CoA生成:在线粒体基质中进行,脱掉2H进入 呼吸链
第六章 线 粒 体
线粒体的形态结构 线粒体的化学组成和酶的分布 线粒体的功能 线粒体的半自主性
第一节 线粒体的形态结构 ⑴
一. 线粒体的基本性质 1894年,Altmann,生命小体(bioblast) 1897年,Benda命名线粒体(mitochondria)
㈠ 形态:光镜下多种多样,有线状,短杆状, 粒状,圆形,哑铃形等。与细胞种类 和生理状态有关。
⑾ DNA ——线粒体的DNA,存在于内腔中。
外膜
核糖体
嵴间腔
内膜
膜间腔
基粒
嵴内 腔
DNA
基质
嵴 基质 颗粒
第二节 线粒体的化学组成和酶的分布⑴
一.化学组成——蛋白质,脂类,水等。 ㈠ 蛋白质:占线粒体干重的65~70%,
内膜含量最多。 可溶性——基质中的酶,外周蛋白
(膜表面) 不溶性——膜镶嵌蛋白,结构蛋白,
形成柄的一部分。
大鼠肝细胞ATP合 成酶的电镜照片
牛心脏细胞基 粒的电镜照片
第一节 线粒体的形态结构 ⑹
⑸ 膜间腔(inter membrane space) ——内、外膜之间的间隙,宽6~8nm,其中有 可溶性酶,底物等。
⑹ 嵴间腔(intercristal space) ——嵴与嵴之间的腔,也称内腔或内室。其内充 满基质。
丙酮酸脱氢酶系
丙酮酸+辅酶A
乙酰CoA
第三节 线粒体的功能 ⑷
㈢ 三羧酸循环:在线粒体基质中进行,脱掉
4×2H进入呼吸链
乙酰CoA+草酰乙酸→柠檬酸→异柠檬酸
↑
↓
苹果酸←延胡索酸←琥珀酸
该过程生成2ATP
第三节 线粒体的功能 ⑸
㈣ 电子传递和氧化磷酸化:在线粒体内膜上 进行 电子传递链(呼吸链)——它是一组酶的 复合体,包括NAD、FMN、FAD、辅酶Q、 细胞色素b、c1、c、a、a3,在电子逐级传 递过程中,完成氧化还原并释放能量。
酶蛋白等
第二节 线粒体的化学组成和酶的分布⑵
㈡ 脂类:占线粒体干重的25~30%,以磷脂为主。 外膜:脂类52%,蛋白质48% 内膜:脂类24%,蛋白质76%
㈢ 水:线粒体中数量最多的一种成分。其功能—— ①酶促反应的溶剂 ②物理介质
㈣ 其它:辅酶Q,NAD,FMN。FAD等成分
第二节 线粒体的化学组成和酶的分布⑶
外膜
核糖体
嵴间腔
内膜
膜间腔
基粒
嵴内 腔
DNA
基质
嵴 基质 颗粒
第一节 线粒体的形态结构 ⑶
二. 线粒体的超微结构 ——电镜下,线粒体是两层单位膜围成的封闭的 囊状结构。
⑴ 外膜(outer membrane) ——位于线粒体外表面,一层单位膜,厚6~7nm, 含有很多运输蛋白,构成小溶质分子可以穿 过的通道。
其实质是ATP酶复合体,是偶联磷酸化的关键装 置。包括:
头部——含可溶性ATP酶(F1),有合成 ATP的功能,其上部有一个抑制多肽。
柄部——是对寡霉素敏感的蛋白(OSCP) 基片——嵌入内膜,为疏水蛋白(HP)
内膜上的特殊结构——基粒(elementary particle)
在内膜和嵴上有许多带柄的小体称为基粒,又称为ATP酶复合体。