无功能垂体腺瘤
无功能垂体腺瘤引发什么疾病

无功能垂体腺瘤引发什么疾病*导读:一、尿崩症大量低比重尿,尿量超过3L/d;因鞍区肿瘤过大或向外扩展者,常有蝶鞍周围神经组织受压表现,如视……*一、尿崩症大量低比重尿,尿量超过3L/d;因鞍区肿瘤过大或向外扩展者,常有蝶鞍周围神经组织受压表现,如视力减退、视野缺失;有渴觉障碍者,可出现脱水、高钠血症、高渗状态、发热、抽搐等,甚至脑血管意外。
*1、实验室检查:(1)尿渗透压:为50~200mOsm/kgH2O,明显低于血浆渗透压,血浆渗透压可高于300mmol/L(正常参考值为280~295mmol/L)。
(2)血浆抗利尿激素值:降低(正常基础值为1~1.5pg/ml),尤其是禁水和滴注高渗盐水时仍不能升高,提示垂体抗利尿激素储备能力降低。
(3)禁水试验:是最常用的有助于诊断垂体性尿崩症的功能试验。
方法:试验前测体重、血压、尿量、尿比重、尿渗透压。
以后每小时排尿,测尿量、尿比重、尿渗透压、体重、血压等,至尿量无变化、尿比重及尿渗透压持续两次不再上升为止。
抽血测定血浆渗透压,并皮下注射抗利尿激素(水剂)5U,每小时再收集尿量,测尿比重、尿渗透压1~2次。
一般需禁水8~12h以上。
如血压有下降、体重减轻3kg以上时,应终止试验。
正常人或精神性烦渴者,禁水后尿量减少,尿比重、尿渗透压升高,故血压、体重常无明显变化,血浆渗透压也不会超过300mmol/L,注射抗利尿激素后尿量不会继续减少,尿比重、尿渗透压不再继续增加。
垂体性尿崩症禁水后尿量减少不明显,尿比重、尿渗透压无明显升高,尤其是完全性垂体性尿崩症,可出现体重和血压明显下降,血浆渗透压升高(大于300mmol/L),注射抗利尿激素后尿量明显减少,尿比重、尿渗透压成倍增高。
部分性垂体性尿崩症变化不如完全性垂体性尿崩症显著,有时与精神性烦渴不易鉴别。
肾性尿崩症患者禁水和肌注抗利尿激素,均不能使尿量减少及尿液浓缩。
(4)颅部及鞍区CT、磁共振检查:有助于该区域器质性病变的诊断和鉴别断。
垂体瘤

鉴别诊断
垂体瘤的诊断应主要与以下几种疾病进行鉴
别:垂体增生、炎症((感染性、非感染性)), 肿瘤、其他。
疾病治疗
垂体瘤治疗目标:抑制肿瘤的自主激素分泌,
最大程度去除肿瘤,维持正常垂体功能,减 轻肿瘤对视力的影响,防止肿瘤复发,防止、 处理并发症。 目前,垂体瘤的治疗方法主要有三种:药物 治疗、手术治疗和放射治疗。
术ቤተ መጻሕፍቲ ባይዱ护理
4、排便指导:术后请勿用力排便,以防止脑腔压力 增高导致脑疝发生,如发生便秘,服缓泻剂或肛注开 塞露等。 5、口腔护理:因口腔内有伤口,所以每日做口腔护 理,保持口腔内的清洁,由于术后用纱条填塞鼻腔止 血,病人只能张口呼吸,易造成口腔干燥,此时应用湿 纱布盖于口唇外,保持口腔湿润,减轻不适,促进食欲。 6、术后补充营养,预防感染:需行静脉输液,输液时 肢体可能感觉麻木,输液后可适当进行局部活动或按 摩。
临床表现
垂体瘤可有一种或几种垂体激素分泌亢进的临床表 现。除此之外,还可能有因肿瘤周围的正常垂体组 织受压和破坏引起不同程度的腺垂体功能减退的表 现;以及肿瘤向鞍外扩展压迫邻近组织结构的表现, 这类症状最为多见,往往为病人就医的主要原因。. 1. 激素分泌过多症候群: 2. 激素分泌减少 3. 垂体周围组织压迫征群 (头痛、视力减退、视野 缺损、 海绵窦综合征、下丘脑综合征、如肿瘤破坏 鞍底可导致脑脊液鼻漏、垂体卒中)
术前护理
术前晚应保证睡眠充足 如有心理紧张难以 入睡者,可依据医嘱给予安定2片口服,并予 以心理疏导。 5 术前禁食、禁水10h 手术前30min予肌注鲁 米那0.1g,阿托品0.5mg。
4
术后护理
饮食的指导对于患者回复健康发挥着很大的
垂体瘤要做那些检查才能确诊

垂体瘤要做那些检查才能确诊绝大多数垂体腺瘤具有较高的分泌功能,使血中激素水平升高,并产生相应的临床症状。
但也有一些垂体腺瘤并不使血中激素水平升高,也无激素过多症状,称为临床无功能垂体腺瘤,简称无功能垂体腺瘤,亦称临床无活性垂体腺瘤(CIPA)、无内分泌活性腺瘤或非分泌性垂体腺瘤,无功能腺瘤占所有垂体腺瘤的25%~30%。
垂体瘤要做那些检查才能确诊?无功能垂体腺瘤的诊断要点为:1.存在垂体瘤的影像学证据。
2.有头痛,视野缺损等垂体占位的表现。
3.无垂体激素过多的临床表现和实验室证据(PRL除外)。
4.有腺垂体功能减退的表现。
5.由于大多数无功能腺瘤患者有血PRL水平的升高,故PRL测定具有重要意义,多数患者血促性腺激素水平降低或在正常范围,但少数患者可有血促性腺激素和(或)其亚单位的升高,性激素的水平一般下降,血TSH,GH及ACTH水平一般正常或轻度降低,其储备功能及靶腺激素水平也多降低,但卓著降低者不多见,偶尔,无功能腺瘤作为亚临床GH瘤或ACTH瘤,则24h尿皮质醇或血IGF-1水平可轻度升高,无功能垂体腺瘤对下丘脑激素的反应具有一定的特点,这在诊断上具有重要意义,常见的利用下丘脑激素的诊断试验有:(1)TRH试验:正常促性腺激素细胞并无TRH受体,故给正常人注射TRH并不引起血LH和FSH水平的升高,大多数无功能腺瘤起源于促性腺激素细胞,约1/3的瘤性促性腺激素细胞含有TRH受体,它们对TRH有反应,约40%的无功能腺瘤病人于注射TRH后血促性腺激素和(或)其亚单位水平升高。
垂体瘤要做那些检查才能确诊?(2)GnRH试验:无功能垂体腺瘤多起源于促性腺激素细胞,这些瘤性促性腺激素细胞含有GnRH受体,故对内源性GnRH,GnRH激动剂性类似物及GnRH拮抗剂都有反应,正常情况下,GnRH对促性腺激素细胞的刺激作用依赖于其特征性脉冲分泌,如连续给予GnRH或长效GnRH类似物则出现失敏现象,促性腺激素分泌反而减少,无功能腺瘤不存在这种失敏现象,这是其特征之一,据Klibanski 等(19)报道,持续给予无功能腺瘤患者长效GnRH类似物DTrp6-Pro9-NEt-LHRH 后,血LH,FSH及α亚单位水平都升高。
无功能垂体腺瘤是怎么回事?

无功能垂体腺瘤是怎么回事?*导读:本文向您详细介绍无功能垂体腺瘤的病理病因,无功能垂体腺瘤主要是由什么原因引起的。
*一、无功能垂体腺瘤病因*一、发病原因无功能垂体腺瘤实际上是一组异质性肿瘤,它们中的大多数具有分裂功能(多为促性腺激素),只是其分泌功能较低,不引起血激素水平的升高,这类肿瘤称为沉寂性腺瘤(silent adenoma)。
有些无功能腺瘤可能确实没有分泌功能,其细胞来源不清,称为裸细胞瘤或无特征细胞腺瘤(null cell adenoma)。
*二、发病机制垂体瘤发病机制的研究曾出现过两种学说,即垂体细胞自身缺陷学说和下丘脑调控失常学说。
现基本统一起来,认为垂体瘤的发展可分为两个阶段起始阶段和促进阶段。
在起始阶段垂体细胞自身缺陷是起病的主要原因,在促进阶段下丘脑调控失常等因素发挥主要作用。
即某一垂体细胞发生突变,导致癌基因激活和(或)抑癌基因的失活,然后在内外因素的促进下单克隆的突变细胞不断增殖,逐渐发展为垂体瘤。
1.垂体瘤细胞自身内在缺陷现在运用分子生物学技术已弄清大多数有功能的及无功能腺瘤是单克隆源性的,源于某一单个突变细胞的无限制增殖。
发生变异的原因为癌基因的激活和(或)抑癌基因的失活。
已查明的主要癌基因有gsp、gip、ras、hst 及PTTG等,抑癌基因有MEN-1、p53、Nm23及CDKN2A等。
其中gsp基因在40%的GH瘤、10%的无功能腺瘤、6% ACTH瘤中发现。
gsp基因及gip2基因的激活使内源性GTP酶活性受到抑制,于是Gs蛋白及Gi2蛋白的 -亚基持续活化,后两者可分别看成是gsp癌基因和gip2癌基因的产物。
这两种癌基因产物可直接引起核转录因子如AP-1、CREB和Pit-l的活化,使激素分泌增多并启动肿瘤生长。
此外,癌基因的激活会导致胞内cAMP水平增加,cAMP可刺激cyclin(细胞周期蛋白)DL和3产生cdk2和cdk4,后两者可促进细胞由G1期进入S期。
垂 体 瘤

垂体瘤一、概述:垂体瘤(The hypophysoma)应包括垂体腺瘤(Pituitary adenoma)、垂体细胞瘤(The Pituicytoma)、垂体腺癌(Malignat adenoma of pituitary)和垂体转移癌(Matastatic carcinomatous of pituitary)等四种病变。
其中垂体腺瘤是颅内最常见的肿瘤,其发病率占颅内肿瘤的第三位,仅次于胶质瘤和脑膜瘤。
国内80年代初期23个医疗单位的统计,垂体腺瘤的相对发生率平均为9.60%(3.89%~16.09%),与国外资料(11.1%)相比较相差不大。
近年来垂体腺瘤的病例有增多的趋势。
其原因除内分泌诊断技术、神经放射等检测技术设备的提高外,跨学科的专业知识的普及,各专业的密切协作及人们的文化素质自我保健意识、生活水平的提高也不无关系。
国外垂体疾病死亡的尸检中,有20%~50%为亚临床的垂体微腺瘤。
因此,目前垂体腺瘤的发病率较80年代的统计为高。
至于垂体细胞瘤,垂体腺癌和垂体转移癌等都是少见的。
二、脑垂体腺瘤【概述】垂体腺是人体重要的内分泌腺体。
垂体腺瘤是内分泌系统肿瘤,是生长缓慢的良性肿瘤。
但近年来对复发性垂体腺瘤的研究认为垂体腺瘤生物学行为有1/3肿瘤具有侵袭性,侵犯海绵窦和周围骨质及硬脑膜,这就造成复发和转移。
H垂体腺瘤术后复发率为10%~36%,因此,事实上的垂体腺瘤难以达到真正的全切除,术后残余是复发的首要因素。
垂体腺瘤多发生于青壮年;小儿及老人少见。
男、女发病率大致相等。
【垂体的解剖】1、垂体为一黄红色横的椭圆形腺体,其横径大约9~12毫米,前后径8~12毫米,垂直径5~8毫米,重0. 5~0.6克,女性稍重些。
脑垂体位于蝶鞍的垂体窝内,上面被硬脑膜形成的鞍隔覆盖,中央有鞍隔孔,是垂体柄通过的地方。
鞍隔有厚薄之分,这决定了垂体腺瘤的发展方向。
垂体的上方为视交叉,两侧有海绵窦颈内动脉。
垂体分为腺垂体(垂体前叶)和神经垂体(垂体后叶)两大部分。
无功能垂体腺瘤怎样治疗?

无功能垂体腺瘤怎样治疗?*导读:本文向您详细介绍无功能垂体腺瘤的治疗方法,治疗无功能垂体腺瘤常用的西医疗法和中医疗法。
无功能垂体腺瘤应该吃什么药。
*无功能垂体腺瘤怎么治疗?*一、西医*1、治疗同其他垂体腺瘤一样,无功能腺瘤的治疗方法有:外科治疗、放射治疗和内科治疗。
目前,仍以外科治疗为首选,手术效果不佳或术后复发者可加用放射治疗,肿瘤压迫症状不著者可试用内科治疗,如内科治疗效果不佳仍应采取手术治疗。
1.外科治疗是否采用外科治疗常取决于肿瘤的大小及临床表现。
对于压迫症状较明显且瘤体较大者一般推荐外科治疗,成功的手术可有效地解除因肿瘤占位效应而产生的一系列症状,而且手术标本可做免疫细胞化学等检查,这对明确诊断具有极为重要的意义。
对于无症状的微腺瘤则并不推荐手术治疗。
一般认为,对于存在视野缺损及神经症状者应尽早手术,以防止视交叉和脑神经出现不可逆性损害。
目前,多采取经蝶术式,此种术式约可使90%的病例视野缺损获得改善,约60%的病例视力可完全恢复。
经蝶手术最主要的并发症是垂体功能减退。
据Arafah报道,术后GH缺乏、甲状腺功能减退、肾上腺皮质功能减退及性腺功能减退的发生率分别为85%、35%、46%和65%。
显然,术后垂体功能减退的发生率显著低于术前,说明经蝶手术在恢复腺垂体功能方面是有效的。
不过,经蝶手术在改善GH 缺乏方面并无很好的效果,原因可能是GH细胞一经损伤就很难恢复。
2.放射治疗手术切除不完全或术后复发者可作术后放射治疗。
有研究表明,术后采用放射治疗者其复发率并不较术后不予放射治疗者低,造成这种结果的原因可能是病例选择上的不同。
目前多数学者认为,术后放射治疗对于提高无功能腺瘤的预后具有一定的意义。
Katznelson等提出,经蝶手术后的第1年内应每6个月随访1次,以后的5年里应每年随访1次,再以后每2年随访1次,每次随访应做MRI及视野检查,如有肿瘤生长的证据应立即开始放射治疗(或考虑再次经蝶手术)。
垂体瘤的治疗方案

一、引言垂体瘤是起源于垂体前叶的肿瘤,可分为功能性垂体瘤和无功能性垂体瘤。
功能性垂体瘤可以分泌多种激素,如生长激素、促肾上腺皮质激素、促甲状腺激素等,导致相应的内分泌症状;而无功能性垂体瘤则不分泌激素,但体积较大时可能压迫周围神经和血管,引起相应的临床症状。
垂体瘤的治疗方法包括药物治疗、手术治疗和放射治疗等。
本文将详细介绍垂体瘤的治疗方案。
二、药物治疗1. 药物治疗的目的(1)控制垂体瘤的生长,减轻肿瘤对周围组织的压迫。
(2)降低激素水平,改善内分泌症状。
(3)为手术治疗创造条件。
2. 药物治疗的方法(1)抑制垂体生长激素分泌的药物:如多巴胺受体激动剂(如溴隐亭)、生长抑素类似物(如奥曲肽)等。
(2)降低激素水平的药物:如糖皮质激素(如泼尼松)、甲状腺激素(如左甲状腺素)等。
(3)抑制垂体分泌促性腺激素的药物:如戈舍瑞林、促性腺激素释放激素类似物(如曲普瑞林)等。
三、手术治疗1. 手术治疗的目的(1)切除垂体瘤,解除肿瘤对周围组织的压迫。
(2)恢复垂体功能,改善内分泌症状。
(3)为药物治疗提供支持。
2. 手术治疗的方法(1)经蝶窦垂体瘤切除术:是目前最常用的垂体瘤切除术式。
通过鼻蝶窦入路,在显微镜下切除肿瘤。
(2)经颅垂体瘤切除术:适用于大型垂体瘤或侵袭性垂体瘤。
(3)放射治疗辅助手术:对于无法完全切除的垂体瘤,可行放射治疗,以缩小肿瘤体积,提高手术切除率。
四、放射治疗1. 放射治疗的目的(1)缩小肿瘤体积,减轻肿瘤对周围组织的压迫。
(2)降低激素水平,改善内分泌症状。
(3)为药物治疗提供支持。
2. 放射治疗的方法(1)立体定向放射治疗:如立体定向放射外科(如伽玛刀、X刀)。
(2)常规放射治疗:适用于体积较大的垂体瘤或药物治疗效果不佳的患者。
五、综合治疗方案1. 早期垂体瘤:对于早期垂体瘤,可根据病情选择药物治疗或手术治疗。
药物治疗适用于内分泌症状较轻的患者,手术治疗适用于肿瘤较大、压迫周围组织或药物治疗效果不佳的患者。
垂体瘤有什么症状

垂体瘤有什么症状*导读:垂体瘤要与原发性垂体瘤相鉴别,是一种发生于垂体前叶的肿瘤,也可发生于垂体后叶或者颅内,颅内转移癌可累及垂体。
垂体瘤有什么症状,下面我们来探讨一下。
……垂体瘤要与原发性垂体瘤相鉴别,是一种发生于垂体前叶的肿瘤,也可发生于垂体后叶或者颅内,颅内转移癌可累及垂体。
垂体瘤有什么症状,下面我们来探讨一下。
1.体征:垂体瘤又叫垂体微腺瘤,常见体征为肢体肥大,无功能肿瘤增大,前者多由激素分泌过多引起,后者多由激素分泌过少引起。
除此之外,闭经、男性不育、阳痿也是垂体瘤的常见症状,主要原因是垂体组织破坏,激素分泌过少。
2.症状:垂体瘤的常见症状为头痛、视力下降、眼底改变、视野不全,此类症状为神经纤维刺激症和视神经束压迫症。
3.检查:垂体瘤的检查方法有气脑和脑血管造影、CT检查、磁共振成像、颅平片检查。
4.治疗:垂体瘤治疗有三大方法,第一是药物治疗法,第二是放射治疗法,第三是手术治疗法。
手术治疗要根据患者诊断劫夺及肿瘤的带下,受压范围和侵蚀情况而定。
治疗不当会引起视力障碍加重、脑膜炎、尿崩症、水、电解质紊乱等并发症。
5.保健:治疗时,患者要注意保健,不要吃辛辣油腻的食物,不要吃煎炸刺激性强的食物,不要吃鲇鱼,要多吃新鲜的蔬菜水果,多吃营养丰富的食物,多吃清淡易消化的食物。
若便秘,可服用通便润肠药;若腹泻,可服用止泻药;若复发,可到医院复查;若体质弱,可经常锻炼提高自身免疫力;若营养不良,可多吃新鲜的蔬菜水果。
垂体瘤有什么症状,相信大家已经知道。
如果大家还有相关疑问,欢迎咨询相关专栏的医生或者各专业医院的医生。
由于垂体瘤对身体危害极大,不仅会生长发育,日常生活质量,而且会影响工作,因此患者发病后要及早治疗,有疑问就多询问医生,这样才能及早恢复健康,免除疾病的困扰,减轻治疗的经济负担,享受美好愉快的生活。
放射外科时代无功能性垂体腺瘤的治疗

放射外科时代无功能性垂体腺瘤的治疗
刘灿虎;张丽君;许谦;金道云
【期刊名称】《国际神经病学神经外科学杂志》
【年(卷),期】2006(33)2
【摘要】无功能性垂体腺瘤因为临床上无明显的内分泌症状,所以发现时往往成为大腺瘤或巨大腺瘤。
开颅手术仍具有一定风险,经蝶窦入路手术切除成了首选治疗方法。
对手术切除后残留的垂体腺瘤放射线治疗仍对患者有好处,立体定向放射外科,特别是伽玛刀治疗获得了高效率抑制肿瘤生长,并且具有对周围正常垂体及神经组织损伤小,精确度高,费时少等优点。
对无功能性垂体腺瘤的治疗主张手术切除与放射线治疗、立体定向放射外科相结合。
【总页数】4页(P177-180)
【关键词】垂体腺瘤;无功能性;手术切除;伽玛刀;放射外科
【作者】刘灿虎;张丽君;许谦;金道云
【作者单位】哈尔滨医科大学附属第二医院;哈尔滨市儿童医院
【正文语种】中文
【中图分类】R736.4
【相关文献】
1.低剂量立体定向放射治疗功能性垂体腺瘤的疗效分析 [J], 洪洋;徐培坤;
2.老年人无功能性垂体腺瘤的显微外科治疗 [J], 闫世军;宋彬;陈岩;朱景华
3.老年无功能性垂体腺瘤的临床特征及显微外科治疗 [J], 赵奕;连伟;幸兵;姚勇;冯
铭;王任直;吴焕文
4.巨大无功能性垂体腺瘤分型及外科治疗方式研究 [J], 望家兴;魏梁锋;谢金华;张帅;林顺安;赵琳;黄银兴;王守森
5.功能性垂体腺瘤放射治疗 [J], 潘绵顺;王鹏;牛朝诗
因版权原因,仅展示原文概要,查看原文内容请购买。
脑垂体瘤的分类

脑垂体瘤的分类
以下是 9 条关于脑垂体瘤分类的内容:
1. 功能性脑垂体瘤知道不?就像身体里的一个活跃小调皮!比如生长激素瘤,它要是捣蛋起来,能让人的外貌变得很不一样呢!你想想,突然变得大手大脚或者额头突出,多奇怪呀!
2. 还有泌乳素瘤呢,这就像是个捣乱的小恶魔!女性要是碰上它,可能月经都不正常了,那可真让人发愁啊,这不是给生活添乱嘛!
3. 促肾上腺皮质激素瘤呀,就如同一个引发混乱的因子!会让身体里的激素乱套,血压呀、血糖呀都可能出问题,这多吓人呀!
4. 无功能性脑垂体瘤呢,就像是个安静的潜伏者!常常没啥明显症状,可到后面发现说不定问题就大啦,你说是不是得注意呀!比如巨大腺瘤,它悄悄地长,等发现可能都晚了呢!
5. 嫌色细胞瘤算得上是比较低调的那种啦,可别小瞧它!就像隐藏在暗处的小麻烦,可能在不知不觉中就带来麻烦啦。
6. 嗜酸性细胞瘤不就是个让人头疼的家伙嘛!稍微不注意,它就可能弄出些状况来,哎呀呀,想想都头大!
7. 嗜碱性细胞瘤呢,好比是个神秘莫测的存在!你永远不知道它啥时候会搞点事情,真让人心里没底呀!
8. 混合型脑垂体瘤啊,那可复杂了,就像是个大杂烩!各种问题交织在一起,处理起来可不容易呢,这可怎么办才好呢?
9. 垂体癌就更别提了,那简直是最可怕的噩梦呀!这可不是开玩笑的,得赶紧重视起来呀!
我觉得脑垂体瘤的这些分类都很重要,了解清楚才能更好地应对呀!。
无功能垂体大腺瘤诊疗进展

无功能垂体大腺瘤诊疗进展吕传奇;廉治刚;董斌;赵永顺;白景阳;陈正昊;李俊【期刊名称】《大连医科大学学报》【年(卷),期】2016(038)006【摘要】垂体无功能腺瘤缺乏典型的临床症状,在临床中很难及时发现,往往因为巨大的垂体腺瘤出现压迫和侵袭症状而被临床医生诊断.无功能垂体大腺瘤的治疗首选手术治疗,根据腺瘤的解剖部位及影像学检查结果,选择最佳的手术方式.由于手术难度大,全切率低,复发率高,通常需要术后辅助相关药物和放射治疗,以期达到最佳治疗效果.本文对无功能垂体大腺瘤的诊断和治疗进展进行综述.【总页数】5页(P611-615)【作者】吕传奇;廉治刚;董斌;赵永顺;白景阳;陈正昊;李俊【作者单位】大连医科大学附属第一医院神经外科,辽宁大连116011;大连医科大学附属第一医院神经外科,辽宁大连116011;大连医科大学附属第一医院神经外科,辽宁大连116011;大连医科大学附属第一医院神经外科,辽宁大连116011;大连医科大学附属第一医院神经外科,辽宁大连116011;大连医科大学附属第一医院神经外科,辽宁大连116011;大连医科大学附属第一医院神经外科,辽宁大连116011【正文语种】中文【中图分类】R651.1+3【相关文献】1.神经导航及内镜辅助下经鼻蝶显微切除无功能性垂体大腺瘤的近期疗效分析 [J], 吴世强;张卓;周明辉;吴亢;陈娟;张华楸;舒凯;雷霆2.经单侧鼻孔蝶窦入路显微手术切除大及巨大无功能垂体腺瘤疗效分析(附66例报告) [J], 黎海滨; 张鹏; 王水娇; 岑庆君3.老年垂体大腺瘤合并医源性库欣综合征、继发性肾上腺皮质功能减退症1例报告与反思 [J], 李佳;陈炯镇;朱凯旋4.老年垂体大腺瘤合并医源性库欣综合征、继发性肾上腺皮质功能减退症1例报告与反思 [J], 李佳;陈炯镇;朱凯旋5.神经内镜联合多模态神经导航系统切除无功能性垂体大腺瘤的临床应用 [J], 呼虹宇;张天鼎;赵君爽;李龙;景治涛因版权原因,仅展示原文概要,查看原文内容请购买。
垂体腺瘤的诊断与治疗进展

垂体腺瘤的诊断与治疗进展垂体腺瘤是一种常见的颅咽管区肿瘤,多数为良性。
它来源于位于脑底部的垂体腺细胞,可以分泌过多或过少的激素。
根据其大小和功能特点,垂体腺瘤可分为微腺瘤和巨腺瘤。
随着影像学技术和外科手术等领域的进步,对于垂体腺瘤的诊断和治疗取得了显著进展。
一、诊断进展:1. 影像学诊断:传统的平片检查已经被功能性磁共振成像(fMRI)所取代。
fMRI可以明确显示肿瘤的大小、位置和影响范围,并且能够检测肿瘤是否侵入周围组织,对于鉴别良性与恶性具有较高准确度。
2. 激素水平检测:血液中相关激素水平异常是垂体腺瘤最主要的诊断指标之一。
通过测量抗利尿激素(ADH)、促甲状腺激素(TSH)、生长激素(GH)和催乳激素(PRL)等的水平,可以辅助判断病变的类型。
3. 病理学检查:对于经影像学检查发现的垂体腺瘤,从手术中获取组织标本进一步行病理学检查是必要的。
光镜下观察肿瘤细胞的形态特点、核分裂象和血管侵犯程度可以帮助判断其恶性程度。
二、治疗进展:1. 药物治疗:对于无功能或压迫性垂体腺瘤,尤其是微小者,药物治疗成为首选。
目前常用的药物有硫酸溴隐亭、多巴胺受体激动剂和类固醇激素等。
这些药物能够控制肿瘤生长和减少相关激素分泌,同时也有利于保护视神经。
2. 外科手术:外科手术是治疗垂体腺瘤最彻底有效的方法之一。
通过经经颅道或经鼻道途径进行手术切除肿瘤,可以缓解患者的症状和改善体征。
目前外科手术已经发展到显微神经导航、内窥镜等技术,使手术更加精准和创伤小。
3. 放射治疗:对于未能手术切除的垂体腺瘤或复发的巨腺瘤,放射治疗是常用的选择之一。
主要包括传统放射治疗和适形调强放射治疗。
它通过高能射线破坏肿瘤细胞,达到控制或减少肿瘤生长的效果。
4. 靶向药物治疗:近年来出现了一些针对特定靶点的抗癌药物,这些药物可以干扰垂体腺瘤细胞中关键信号通路,从而实现对肿瘤细胞增殖和分化的抑制。
综上所述,随着医学技术的不断进步,在垂体腺瘤的诊断和治疗方面取得了显著进展。
脑垂体瘤
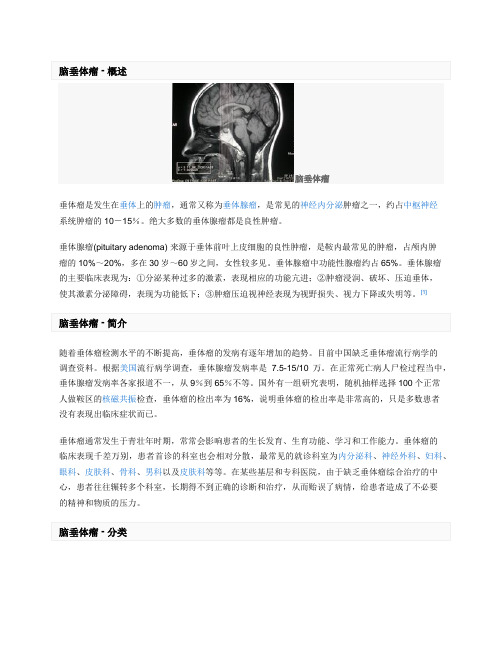
脑垂体瘤垂体瘤是发生在垂体上的肿瘤,通常又称为垂体腺瘤,是常见的神经内分泌肿瘤之一,约占中枢神经系统肿瘤的10-15%。
绝大多数的垂体腺瘤都是良性肿瘤。
垂体腺瘤(pituitary adenoma) 来源于垂体前叶上皮细胞的良性肿瘤,是鞍内最常见的肿瘤,占颅内肿瘤的10%~20%,多在30岁~60岁之间,女性较多见。
垂体腺瘤中功能性腺瘤约占65%。
垂体腺瘤的主要临床表现为:①分泌某种过多的激素,表现相应的功能亢进;②肿瘤浸润、破坏、压迫垂体,使其激素分泌障碍,表现为功能低下;③肿瘤压迫视神经表现为视野损失、视力下降或失明等。
[1]随着垂体瘤检测水平的不断提高,垂体瘤的发病有逐年增加的趋势。
目前中国缺乏垂体瘤流行病学的调查资料。
根据美国流行病学调查,垂体腺瘤发病率是7.5-15/10万。
在正常死亡病人尸检过程当中,垂体腺瘤发病率各家报道不一,从9%到65%不等。
国外有一组研究表明,随机抽样选择100个正常人做鞍区的核磁共振检查,垂体瘤的检出率为16%,说明垂体瘤的检出率是非常高的,只是多数患者没有表现出临床症状而已。
垂体瘤通常发生于青壮年时期,常常会影响患者的生长发育、生育功能、学习和工作能力。
垂体瘤的临床表现千差万别,患者首诊的科室也会相对分散,最常见的就诊科室为内分泌科、神经外科、妇科、眼科、皮肤科、骨科、男科以及皮肤科等等。
在某些基层和专科医院,由于缺乏垂体瘤综合治疗的中心,患者往往辗转多个科室,长期得不到正确的诊断和治疗,从而贻误了病情,给患者造成了不必要的精神和物质的压力。
脑垂体瘤垂体瘤可以按照肿瘤的大小和激素分泌的功能不同来分类。
根据肿瘤大小的不同,垂体瘤分为垂体微腺瘤(肿瘤的直径小于1cm)和垂体腺瘤(肿瘤直径大于等于1cm)。
根据分泌激素的不同,又可以分为激素分泌性垂体瘤和无功能腺瘤。
垂体肿瘤的大小与治疗预后关系密切。
大约70%以上的垂体无功能瘤是不需要治疗的,若没有相应的垂体功能低减的情况,临床上可以长期随诊。
垂体腺瘤概述及治疗

垂体的解剖及生理学
(Anatomy and physiology)
(1)解剖及生理
脑下垂体位于蝶鞍内。呈卵圆形,约大小,平均重量750mg(男350-700mg,女450-990)。女性妊娠时呈生理性肥大。
01
腺垂体由外胚层的拉克氏囊(Rathke氏)囊分化而来,神经垂体来自前脑底部的 神经外胚层。垂体藉垂体柄与三脑室底和侧壁的下丘脑有密切的联系。垂体前叶分成漏斗部,包围着垂体柄(鞍上)和远侧部(鞍内)
垂体激素过量分泌引起一系列的代谢紊乱和脏器损害 肿瘤压迫使其他垂体激素低下,引起相应靶腺功能低下 压迫蝶鞍区结构,如视交叉、视神经、海绵窦、脑底动脉、下丘脑、第三脑室、甚至累及额叶、颞叶、脑干等,导致相应功能的严重障碍。 垂体瘤好发于青壮年,对病人生长、发育、劳动能力、生育功能有严重损害,并造成一系列社会心理影响。患有垂体瘤疾病后应积极早期治疗。
肉芽肿:症状近似垂体脓肿,而且发展更快,除头痛,视力减退外,经常发生多组颅神经受损,多伴有垂体功能低下.病变累及范围广泛,例如鞍内,蝶窦内,鞍上,前中后颅等部位.影像学表现病变周遍硬膜强化明显。
胞性垂体炎:尿崩为主要临床表现。部分伴有垂体功能低下。影像学表现垂体柄明显增粗。垂体组织不同程度地增大。
垂体腺瘤的分类
1
既往按肿瘤细胞染色特性——嫌色性、嗜酸性、嗜碱性
2
目前为细胞分泌功能分类法——泌乳素腺瘤(PRL瘤)、生长激素腺瘤(GH瘤)、促肾上腺皮质激素瘤(ACTH瘤)等,有混合型腺瘤。
如肿瘤直径分—— <1cm,限于鞍内生长称微腺瘤,确诊靠CT或MRI及血清激素水平测定,肿瘤直径 >1cm,突破鞍膈为大腺瘤,>3cm 为巨腺瘤。
素细胞腺瘤:
有关垂体腺瘤的几个问题

有关垂体腺瘤的几个问题出率垂体腺瘤的发生率约10~%(均匀 %)。
~15/10 万人,尸检发现率约~84%(均匀%)。
正常人群随机MRI检一、初期诊疗垂体腺瘤的临床特色是内分泌改变和视力阻碍。
微腺瘤(<1cm)限制于鞍内,初期会出现相应的内分泌征象,应用放射免疫测定法,能正确查明血液激素,有益于垂体腺瘤的初期诊疗,待肿瘤增大向鞍上扩展,压迫视神经、视交错,才出现双颞侧偏盲和视力减退。
1.内分泌改变:垂体腺瘤按内分泌功能分为无功能腺瘤和有功能腺瘤,后者包含泌乳素( prolactin ,PRL)腺瘤、生长激素(growth hormone , GH)腺瘤、促肾上腺皮质腺瘤(adrenocorticotropic hornome , ACTH)腺瘤、促甲状腺(thyroid stimulating hormone , TSH)腺瘤、促性腺激素( gonadotripic hormone , GnH)腺瘤, GnH 腺瘤又包含卵泡刺激素(follicular stimulating hormone ,FSH)和黄体生成素(luteinizing hormone ,LH)腺瘤。
PRL腺瘤( 50%~60%)、GH腺瘤( 20%~30%)及 ACIH腺瘤( 5%~15%)最常有,临床各具不同的内分泌改变。
PRL腺瘤表现为泌乳、闭经、不孕三连症(发育后); GH 腺瘤表现为巨人症(发育前)、肢端肥大症(发育后),巨人伴肢端肥大症(发育前后);ACTH腺瘤变现为向心性肥胖(Cushing’s disease)。
2.视力视线阻碍:增大的垂体腺瘤直接压迫视交错惹起双颞侧偏盲。
微腺瘤未直接压迫视交错,但视交错也可有功能阻碍,表现为典型的双颞侧偏盲,其原由在于高灌输状态下的垂体增殖性病变,经过它对与视交错共同供给血管分支的“盗血” 或扰乱其正常血供,使视交错中部存在的微循环单薄环节发生血供阻碍,而致使颞侧视线缺损。
女性垂体瘤的症状有哪些呢

当今社会很多人对于肿瘤的疾病越来越关注,因为它会直接影响患者的身体健康,所以对于脑瘤的问题都在想办法进行改善。
随着社会发展,脑瘤疾病中发病率最高的就是垂体瘤的问题。
垂体瘤约占颅内肿瘤的10%。
垂体瘤大部分为良性腺瘤,极少数为癌。
虽然不会直接威胁患者生命,但是也会给患者健康带来影响,尤其是女性。
那么,女性垂体瘤的症状有哪些呢?临床专家介绍说,垂体瘤是一组从垂体前叶和后叶及颅咽管上皮残余细胞发生的肿瘤。
此组肿瘤以前叶的腺瘤占大多数,来自后叶者少见。
据不完全统计,其中PRL瘤最常见约占50-55%,其次为GH瘤20-23%,ACTH瘤5-8%,TSH瘤与LH/FSH瘤较少见。
无功能垂体腺瘤,占20-25%。
那么,女性垂体瘤的症状有哪些呢?女性垂体瘤的症状:1、垂体功能低下垂体功能低下的程度与正常垂体腺细胞受压的程度有关。
由于垂体前叶机能低下以致其所控制的内分泌腺萎缩表现多方面的功能障碍。
垂体嫌色性腺瘤或嗜酸性腺瘤的晚期常表现为垂体功能低下。
2、垂体功能亢进嗜酸性细胞能产生生长激素及催乳激素。
嗜碱性细胞产生促甲状腺激素、促肾上腺激素及促性腺激素等。
因此 这类肿瘤可造成垂体功能亢进 尤其是在早期。
嗜酸性腺瘤虽可有生长激素分泌过多 但又可挤压嗜碱性细胞 造成其功能低下。
3、垂体后叶及丘脑下部受累症状肿瘤影响垂体后叶或丘脑下部则可产生嗜唾、多饮、多尿、体温低下以及水、电解质及脂肪代谢障碍等。
4、视神经受压症状与体征由于肿瘤生长在蝶鞍内 向上发展则压迫视交叉或视神经而造成视力、视野的改变 90% 以上的病人都有视力减退 也可为单眼视力减退 甚至造成一目或双目失明。
视野的改变可因压迫神经或视交叉而有所不同 视交叉受压则产生双颞侧偏盲 或一眼颞盲 肿瘤如位于一侧压迫视束或视神经时 可产生对侧同向偏盲 或一眼全盲。
5、内分泌及代谢障碍症状及体征垂体腺内分泌功能很复杂 主要是在中枢神经的支配下 通过内分泌控制着人体的生长、发育、物质代谢及性器官、性功能等生理活动的调节。
无功能垂体腺瘤应该如何预防?

无功能垂体腺瘤应该如何预防?
*导读:本文向您详细介绍无功能垂体腺瘤应该如何预防,
常见预防措施有哪些。
以及无功能垂体腺瘤应该如何护理,无功能垂体腺瘤常见的护理办法有哪些等方面内容。
*怎样预防无功能垂体腺瘤:
*一、预防
在一些体积特别大的垂体无功能瘤,手术前有时需要做血管造影检查,以判明肿瘤与血管之间的关系,避免在手术中损伤血管,发生意外。
*以上是对于怎样预防无功能垂体腺瘤方面内容的相关叙述,那么,下面再看下无功能垂体腺瘤的护理方法,无功能垂体腺瘤的常见护理措施。
*无功能垂体腺瘤常见护理方法:
*一、护理
1、调整日常生活与工作量,有规律地进行活动和锻炼,避
免劳累。
2、保持情绪稳定,避免情绪激动和紧张。
3、保持大便通畅,避免用力大便,多食水果及高纤维素食物。
4、
避免寒冷刺激,注意保暖。
*温馨提示:以上就是对于怎样预防无功能垂体腺瘤,无功能垂体腺瘤的护理方法方面内容的介绍,更多相关资料请继续关注疾病库,或者在站内搜索“无功能垂体腺瘤”可以找到更多,希望以上内容对您有帮助!。
脑垂体腺瘤是什么了解这种常见的颅内肿瘤

实验室检查:激素 水平异常,如生长 激素、促肾上腺皮 质激素等
病理学诊断:金标 准,可明确诊断及 分型
诊断流程
病史采集:了解患 者的症状、体征及 家族史
影像学检查:如 MRI、CT等,观 察脑垂体腺瘤的大 小、位置及与周围 组织的毗邻关系
实验室检查:测定 激素水平,如生长 激素、促甲状腺激 素等,有助于判断 腺瘤的性质
脑垂体腺瘤
,
汇报人:
目录
01 脑 垂 体 腺 瘤 的 定 义
02 脑 垂 体 腺 瘤 的 症 状
03 脑 垂 体 腺 瘤 的 诊 断
04 脑 垂 体 腺 瘤 的 治 疗
05 脑 垂 体 腺 瘤 的 预 防垂体腺瘤的概念
脑垂体腺瘤是一 种起源于脑垂体
的肿瘤
脑垂体腺瘤可以 分为功能性腺瘤 和无功能性腺瘤
预防措施的成本效益:评估预防措施的经济成本和效益,以确定其经济可 行性。 预防措施的长期效果:评估预防措施的长期效果,以了解其可持续性和稳 定性。
预防的长期性
定期体检:定期 进行头部影像学 检查,以便早期 发现脑垂体腺瘤
健康饮食:保持 均衡的饮食,多 吃蔬菜水果,减 少高热量、高脂
肪食物的摄入
控制情绪:保持 心情舒畅,避免 过度紧张和焦虑
坚持运动:适当 的运动有助于提 高身体免疫力, 预防脑垂体腺瘤
的发生
添加标题
添加标题
添加标题
添加标题
THANKS
汇报人:
添加 标题
添加 标题
视力视野障碍:肿瘤压迫视神经导致视力下降和视野缺损
激素分泌异常:如生长激素、促肾上腺皮质激素、促甲状腺激素等分泌过多或过少,导 致相应症状
添加 标题
脑脊液循环障碍:脑垂体腺瘤可能压迫脑脊液循环通路,导致颅内压进一步升高
得了垂体瘤怎么办

得了垂体瘤怎么办人体大脑中的垂体组织主要负责控制人体神经、内分泌系统,垂体瘤即垂体组织出现瘤体,一般这种肿瘤基本都是良性,但也有少数得了垂体瘤的患者会被诊断出恶性病变。
通常情况下,得了垂体瘤,患者需要调整心态,不要惊慌,保持情绪稳定的去主动了解该病病理特征,认识该病并非不治之症,然后第一时间与医生沟通交流,掌握自身垂体瘤性质、类型、病情程度等信息,积极配合医生,采取最佳治疗方案,谨遵医嘱行事,都可取得良好的治疗预后效果。
本文将对垂体瘤相关医学知识进行全面科普分析,以期让更多人能够了解该疾病,从而养成良好的防治意识。
1、垂体瘤类型特征1.1分泌激素垂体瘤临床对垂体瘤分类,大致可分为分泌激素的垂体瘤和不分泌激素的垂体瘤,也称之为垂体有功能腺瘤和垂体无功能腺瘤。
其中分泌激素的垂体瘤,通俗来讲,就是其会分泌大量激素,导致人体内分泌系统受损,从而对患者生命健康产生较大威胁。
比如泌乳素瘤便是一种常见的分泌激素垂体瘤,乳素过量分泌,致使女性月经周期失调,甚至闭经,尤其像青年女性群体,一旦患上泌乳素瘤,无法受孕的风险也会因此而大幅升高;而像老年女性患者群体,骨质疏松风险发生率也会全面增加,对于男性患者群体而言,则多有性欲减退、性功能障碍等表现。
1.2不分泌激素垂体瘤不分泌激素垂体瘤即垂体无功能腺瘤,通俗来讲该类垂体瘤即不会对人体产生任何有作用的激素,反而会压迫正常垂体组织,导致正常垂体组织受压出现功能障碍等现象,最终便会引发人体激素分泌大幅减低的状况发生,所以不分泌激素垂体瘤可看作是分泌激素垂体瘤的“反面”,一个是过量分泌激素、一个是过少甚至不分泌激素。
大部分情况下,不分泌激素垂体瘤基本都处于低功能状态,随时间推移,肿瘤持续增加,压迫垂体周围正常结构,患者便会有明显的头痛、视野缺失等临床征象,即使不分泌激素垂体瘤较小,占位有限,同样也会对人体健康,甚至生命安全产生较大威胁,一经确诊必须尽快予以临床干预治疗,谨防患者病情持续加重的状况发生。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
【疾病名】无功能垂体腺瘤【英文名】nonfunctioning pituitary adenoma【别名】clinically inactive pituitary adenoma;endocrineinactive adenoma;functionless pituitary adenoma;nonsecretory pituitary adenoma;非分泌性垂体腺瘤;临床临床无活性垂体腺瘤;无机能垂体腺瘤;无内分泌活性腺瘤;nonfunctioning hypophyseal adenoma【ICD号】D35.2【病因和发病机制研究的进展】1.病因研究进展2.发病机制研究进展(1)分子生物学发病机制:分子遗传学研究表明:无功能性垂体腺瘤是单克隆起源,其中少部分肿瘤表现为常染色体显性症状,即多发性内分泌肿瘤Ⅰ型(MEN1),其与肿瘤抑制基因MEN1的突变有关;其他肿瘤与染色体11q13的杂合子丢失相关。
30%的生长激素腺瘤发生Gas基因的显性突变,但该突变在其他垂体腺瘤中发生率较低。
有研究表明:在无功能性垂体腺瘤中,可有视黄醇类X 受体、雌激素受体和甲状腺激素受体的下降,这可能与激素的调控有关。
然而,这些因素与垂体腺瘤之间的关系仍不明确。
表皮生长因子受体在80%的无功能性垂体腺瘤中过度表达,而在功能性垂体腺瘤中不表达;在体外,表皮生长因子可促进无功能性垂体腺瘤的生长,而无功能性垂体腺瘤可上调表皮生长因子mRNA。
(2)MEG3a和MEG3基因的发现:为了明确垂体腺瘤的分子生物学发病机制,Zhang等利用cDNA-RDA方法比较正常垂体组织和无功能性垂体腺瘤间基因表达的差异。
他们克隆出一cDNA,该cDNA在无功能性垂体腺瘤中不表达,是已被发现但功能未知的母本印记基因MEG3的新型转录产物;印记基因MEG3在正常人促性腺激素细胞中有表达,而在促性腺激素细胞起源的无功能性垂体腺瘤中不表达;Northern Blot和RT-PCR分析进一步表明:MEG3在功能性垂体腺瘤及许多癌细胞株中也不表达。
此外,MEG3的异位表达能抑制许多癌细胞株的增殖,包括Hela细胞、MCF-7和H4组蛋白等。
基因分析表明:MEG3定位于染色体14q32.3。
Miyoshi等于2000年首次发现MEG3基因是一小鼠基因Gtl2的人类同系物。
众所周知,小鼠12号染色体远端的父系副本可致晚期胚胎死亡并促进细胞生长,而母系副本可致晚期胚胎死亡且抑制生长。
人类14号染色体的父系或母系单亲二倍体与小鼠12号染色体远端是相似的,已有报道表明:该区域对生长、智力活动和肌肉骨骼的多态性有一定的印记效应。
为了分离该区域的印记基因,Miyoshi等从小鼠MEG基因中分离出7个候选克隆产物,其中之一就是MEG3,其与Gtl2基因相同。
由于小鼠MEG3或Gtl2基因和人类MEG3均无明显的开放读码框,故这些基因的功能尚不清楚。
最近报道12号染色体有插入突变的转基因小鼠Gtl2L acZ,当其转基因来自父系时可表现为胎儿期和出生后的生长迟缓;随后,S chuster-Gossler等从转基因整合位点附近分离出Gtl2基因,认为其是父系等位基因的特异性表达产物。
小鼠MEG或Gtl2基因和人类MEG3基因是分别在小鼠12号染色体远端和人类第14号染色体长臂发现的首例印记基因。
MEG3或Gtl2的功能尚不明确,其大转录产物为7kb,有开放读码框,编码产物可能为一177氨基酸的蛋白质;然而在起始ATG附近并无K o z a k 共有序列,且与已知蛋白间无同源性。
S chuster-Gossler等已阐明小转录产物(1.9kb)和其他转录产物的开放读码框更小,也无K o z a k共有序列。
研究Gtl2和MEG3基因的功能,鉴定小鼠12号染色体和人类14号染色体长臂邻近的印记基因,对于研究基因疾病的发病原理和印记基因的作用机制均有极大帮助。
随后几个研究小组报道Gtl2或MEG3基因与另一个印记基因Dl k1密切相关。
Dl k1编码包含6个表皮生长因子重复序列的跨膜蛋白,但Glt2或MEG3基因的功能还不明确。
(3)MEG3a和MEG3的差异:Zhang等在利用cDNA-RDA技术比较分析了正常垂体组织和垂体腺瘤,发现并克隆了MEG3acDNA片段,其在不同组织中表达亦不相同,在正常垂体组织中表达高,而在无功能性垂体肿瘤中低表达或不表达。
MEG3a是MEG3的一种亚型,用该cDNA片段去筛选一人胎儿肝脏基因库,得到的阳性克隆产物再进行亚克隆,并分析其DNA序列。
所有阳性cDNA克隆产物均含有与先前报道MEG3相似的DNA序列,然而并非完全相同。
因为MEG3a在MEG3cDNA中间插入一段141b p的DNA序列。
他们将此特殊的cDNA称为MEG3a,其序列已被提交到GenBan k(登记号为A Y314975)。
MEG3和MEG3a的cDNA序列中均含两个开放读码框,但两者均无一致的K o z a k序列。
在菌落形成和生长速率分析中,MEG3a能极大抑制细胞生长和分化,即使是恶性程度很高且生长迅速的恶性肿瘤细胞,例如宫颈癌的海拉细胞株,乳腺癌的MCF-7及神经胶质瘤H4。
但目前还不明确,该基因的生长抑制作用是受RNA转录产物介导或未知翻译蛋白质的调控;尚无直接证据表明:MEG3a能调控垂体腺瘤细胞的生长,或MEG3a表达缺失会导致垂体肿瘤的形成。
然而,MEG3a能极大抑制细胞生长,且在正常促性腺激素细胞中表达,而在促性腺细胞来源的无功能性垂体腺瘤中不表达,均表明它在维持正常促性腺细胞生长中意义重大。
如果能够比较MEG3和MEG3a,对了解MEG3基因与肿瘤形成间的关系意义深远。
因为MEG3a在正常垂体细胞和成纤维细胞中高表达,而在垂体肿瘤和大多数癌细胞中不表达,所以我们推断MEG3的生物学功能与控制细胞增殖有关。
(4)MEG3a和MEG3基因的组织分布:Zhang等为了解从E S T数据库获得的MEG3与人DNA基因库筛选得到的MEG3a之间的差别,他们把这些cDNA序列与人类基因组列相比较。
通过扩展搜索人类基因组序列,得出MEG3基因定位于染色体14q32.3。
比较MEG3 cDNA亚型序列和该区域的人类基因组序列,得出了MEG3的基因组结构。
MEG3a和MEG3序列的区别在于MEG3a中插入了一段141b p 的DNA片段,而这是由于MEG3a cDNA使用外显子5所造成。
在人类基因组中,我们只发现一段高度匹配的基因序列,提示MEG3是单拷贝基因。
Zhang等同时利用MEG3 cDNA N'末端一段200b p序列作为探针检测其在不同组织中的分布情况,表明MEG3在垂体和小脑中高表达,大脑的其他区域也有表达;同时,胎盘、肾上腺、胰腺和子宫中也有MEG3的表达。
Miyoshi等利用小鼠MEG3或Gtl2序列在d b E S T中进行扩展查询,发现几个匹配度较高的人类E S T克隆产物,这些E S T克隆产物已被绘制到人类14号染色体长臂上。
随后他们对4个E S T克隆产物进行基因序列分析,确定了人类MEG3基因的cDNA结构为一1574b p的转录产物;除了与小鼠外显子4和外显子6,7,8相对应的序列外,其与小鼠MEG3或Gtl2具有极高的同源性(67%),其中前者在人类MEG3序列中缺失,而后者则被完全不同的序列所替代。
同时也发现在成年人中,人MEG3在脑组织中表达,而在其他组织如心脏、肾脏、脾脏、肝脏和结肠中不表达。
(5)MEG3a和MEG3基因缺失的机制:Zhao等研究了人类无功能性垂体腺瘤中MEG3基因表达缺失的潜在机制。
他们在被检肿瘤中未发现基因组的异常,并提出表观遗传学的改变与MEG3基因的表达缺失密切相关。
通过与正常垂体组织比较,他们在不表达MEG3的垂体肿瘤中发现,在第一个外显子的前方及其上游大约1.6~2.1kb内的2段5'侧翼区发生了超甲基化;而且,用甲基化抑制剂处理后能够恢复人类癌细胞株表达MEG3。
因此,他们得出结论认为:MEG3基因调控区的超甲基化与无功能垂体腺瘤中MEG3基因表达的缺失密切相关。
【诊断研究进展】1.辅助诊断检查进展(1)实验室检查进展:①垂体肾上腺轴功能测定:垂体无功能瘤发生继发性肾上腺皮质功能减退的患者表现为ACTH和皮质醇水平下降,对CRH兴奋试验无反应。
垂体肾上腺轴功能测定不仅可以了解患者肾上腺功能状态,特别是对需要作垂体手术的患者,可以及时补充糖皮质激素,避免手术中发生危险。
②垂体甲状腺功能测定:垂体无功能瘤伴有甲状腺功能减退的患者T S H及甲状腺激素水平常低于正常,TRH兴奋试验中,T S H及甲状腺激素水平无明显升高。
③垂体性腺轴功能测定:垂体无功能瘤引起垂体功能减退时,垂体-性腺轴常常最早受到累及,约50%的患者可以发现其F S H、L H及T(E2)基础水平下降,促性腺激素释放激素兴奋试验时约50%患者可发现垂体对GnRH反应低下,不出现F S H、L H及性激素水平的升高。
但是,在少数病例,也可能观察到F S H、L H 水平上升的现象,目前认为这种情况与激素测定中使用的F S H、L H抗体与这些糖蛋白激素的亚单位存在交叉反应有关。
④生长激素测定:生长激素缺乏在垂体无功能瘤患者也较常发生,但由于成人的生长激素不高,且受睡眠等因素的影响,单纯生长激素测定并无太大的意义。
胰岛素或生长激素释放激素激发试验对判定生长激素分泌状态的意义较大,约80%垂体无功能瘤患者在作胰岛素低血糖激发试验及GHRH激发试验时,缺乏生长激素升高的反应。
⑤催乳素测定:约50%垂体无功能瘤病例伴有高催乳素血症,但如与催乳素瘤比较则相对处于较低水平,很少超过100µg/L。
如果催乳素水平高于100µg/L则提示垂体催乳素瘤可能。
⑥α亚单位测定:垂体激素F S H、L H和T S H都由相同的α-亚基及不同的β-亚基所构成,不同的激素功能主要表现在β-亚基。
α亚单位测定以及α亚单位与T S H、F S H、L H等糖蛋白激素的比例可以作为诊断分泌糖蛋白激素的垂体瘤的参考指标。
如α亚单位水平升高,应考虑垂体糖蛋白激素腺瘤或垂体α亚单位瘤的可能,在垂体无功能瘤的鉴别诊断中有一定价值。
(2)特殊检查进展:①影像学检查:由于垂体无功能瘤一般体积较大,有时仅作头颅的正侧位X线摄片也能发现蝶鞍在影像学上存在异常,如蝶鞍扩大、骨质破坏、双鞍底影等,可作为诊断垂体无功能瘤的参考。
更常用的方法是CT和MR I,诊断垂体肿瘤灵敏度较高。
MR I显示垂体肿瘤与周围组织的关系较CT更清晰。
对某些体积特别大的垂体无功能瘤,术前有时需要作血管造影检查,以明确肿瘤与血管的关系,避免术中损伤血管。