基因工程_第八章_基因的表达
08 第八章 基因工程与体外表达

真核生物基因的结构特点
真核生物结构基因示意图
69
原核生物基因的结构特点
基因是连续的, 无内含子
70
一、大肠杆菌表达系统
大肠杆菌表达外源基因的优势 繁殖迅速 培养简单 操作方便 遗传稳定
71
大肠杆菌表达外源基因的劣势
1.缺乏对真核生物蛋白质的修饰加工系统 如糖基化或磷酸化等修饰作用
2. 内源性蛋白酶降解空间构象不正确的异源蛋白,
造成表达产物不稳定
72
(一)基因表达的基本要素 1.目的基因和密码子 (1)目的基因 目的基因如果来自真核细胞必须是 cDNA 。 cDNA的始密码子(ATG)上游部 分(5ˊ端非编码区)必须除去。对于一些分 泌性蛋白,还应除去信号肽部分。
73
(2)密码子 生物体对密码子的偏爱性 密码子偏爱性对外源基因表达的影响
54
因此,当载体的多克隆位点没有插入外
源DNA片段时,α互补能正常进行,产生有活 性的β-半乳糖苷酶。 能使人工底物X-gal变 成蓝色, 产生蓝色菌斑或菌落。如果载体的多 克隆位点插入了外源DNA片段,导致产生无α 互补能力的氨基端片段, 带重组体的细菌形成 白色菌斑或菌落。
55
(二) 免疫学方法
(2) 插入片段方向性的鉴定
60
第四节
真核细胞转染
一. 真核细胞转染的方法与基本原理
(一)磷酸钙沉淀法
(二)电穿孔法 (三)DEAE-葡聚糖法 (四)脂质体介导的基因导入
(五)显微注射法
61
二. 转染细胞的筛选
( 一) TK-细胞突变株筛选转染细胞 细胞内TTP的合成有两条途径 从头合成 可被氨甲喋呤阻断 补救合成 胸苷激酶(TK)是关键酶 TK+: 细胞生长 TK- 细胞 + 含TK基因的载体: 细胞生长
《生物化学》-第八章

➢ 与前述操纵子的基本组成一样,乳糖操纵子也是由结构基因和调控区组成的 ➢ 乳糖操纵子包括Z、Y和A三个结构基因 ➢ Z结构基因编码β-半乳糖苷酶,催化乳糖转变为别乳糖 ➢ Y结构基因编码半乳糖透过酶,促使半乳糖透过酶进入细菌内 ➢ A结构基因编码乙酰转移酶,催化半乳糖形成乙酰半乳糖 ➢ 调控区包括调节基因(I)、启动子(P)、操纵基因(O)及启动子上游的一个CAP结合位点,
第一节 基因表达的调控
二、基因表达调控的概念和意义
(一)基因表达调控的概念
➢ 基因表达调控是指细胞或生物体在接收内外环境信号刺激 或适应环境变化的过程中,在基因表达水平上所做出的应 答,即基因组内的基因如何被表达、表达多少等
➢ 基因表达调控大致可以在5个层次上进行,即转录前、转 录、转录后、翻译和翻译后
➢ 基因表达是指在一定的调节机制的控制下,基因组DNA经 转录、翻译等一系列过程,合成具有特异生物学功能的蛋 白质的过程
➢ 并非所有基因表达过程都产生蛋白质,rRNA、tRNA编码 基因转录生成功能型RNA的过程也属于基因表达
第一节 基因表达的调控
一、基因表达的概念、特点及方式
(二)基因表达的特点--时间特异性
5′-侧上游,主要控制整个结构基因群的转录
第一节 基因表达的调控
三、原核生物基因表达的调控
(一)操纵子的基本组成
➢ 3.操纵基因 ➢ 操纵基因是指能被阻遏蛋白特异性识别并结合
的一段DNA序列,常与启动子邻近或与启动子 序列重叠 ➢ 当阻遏蛋白结合在操纵基因上,阻遏蛋白会阻 碍RNA聚合酶与启动子结合或使RNA聚合酶 不能沿DNA链向前移动,从而阻遏转录的进行
(一)操纵子的基本组成
➢ 1.结构基因 ➢ 操纵子中被调控的编码蛋白质的基因称为结构基因 ➢ 一个操纵子中含有2个以上的结构基因,多的可达20个以上 ➢ 各结构基因头尾衔接、串联排列,组成结构基因群
《目的基因的表达》PPT课件

➢ 包含体表达的优点
o 可以避免宿主的降解。 o 当以包含体表达时,目的基因产物的表达量也往往比较高, o 包含体容易纯化,往往仅通过简单的差速离心及洗涤等几步即可得到较
高纯度的目的蛋白。 o 当所表达的重组蛋白产物对宿主细胞具有毒性时,使重组目的产物以无
活性的包含体形式表达可能是蛋白表达的最佳方式。
一、大肠杆菌表达系统的特点 二、大肠杆菌表达融合蛋白 三、大肠杆菌细胞包含体 四、提高克隆基因在大肠杆菌表达效率的途径
为了在大肠杆菌中合成某种特殊的真核生物的蛋白质以满足商品生产 的广泛需求,仅仅停留在检测水平上的表达是远远不够的,所以,必须设 法提高克隆基因的表达效率。
许多因素诸如启动子的强度、DNA转录起始序列、密码子的选择、 mRNA分子的二级结构、转录的终止、质粒的拷贝数以及质粒的稳定性和 寄主细胞的生理特征等,都会不同程度地影响到克隆基因的表达效率,因 而从分析这些因素入手就能够寻找提高克隆基因表达效率的有效途径。
克隆基因末端的转录终止区的存在,可避免以下几种不利现象:
①若干非必须的转录本的合成,会使细胞消耗巨大的能量用于制造大 量非必须的蛋白质;
②在转录本上有可能形成一些不期望其出现的二级结构,从而降低了 转译的效率;
③偶然会出现启动子阻塞现象,即克隆基因启动子所开始的转录干扰 另一个必要的基因或调节基因的转译。
第八章 目的基因的表达
第一节 基因工程中的基因表达
一、制约目的基因表达的因素 二、阅读框架 三、启动子的影响 四、终止子的影响 五、翻译过程对表达的影响
不论基因工程的研究目的如何,最终均涉及到目的基因的表达问 题。
绝大多数的基因工程研究是以获得基因表达产物为目标。在构建重 组体DNA分子和选择宿主细胞时,还须考虑外源基因表达的问题。
基因工程中的基因克隆与基因表达实验总结

基因工程中的基因克隆与基因表达实验总结基因工程作为一门新兴的交叉学科,已经广泛应用于生物医学、农业、环境保护等领域。
其中,基因克隆和基因表达实验是基因工程的核心技术,对于研究基因功能和开发新药已经起到了重要作用。
本文将对基因工程中的基因克隆和基因表达实验进行总结,并探讨其在科学研究和应用中的前景。
一、基因克隆实验基因克隆是通过重组DNA技术,将感兴趣的基因从一个生物体中复制并插入到另一个生物体中的过程。
它是研究基因功能、生物制药和转基因等领域的基础。
基因克隆实验主要包括以下几个步骤:1. DNA提取与限制性内切酶切割:通过提取DNA样品,使用限制性内切酶切割将目标基因和载体DNA切割成相应片段。
2. 基因插入:将目标基因与载体DNA片段进行连接,常用的方法是使用DNA连接酶将两者黏合。
3. 转化与筛选:将连接后的DNA转入到宿主细胞中,使其成为转基因细胞。
通过选择性培养基进行筛选,可以获得拥有目标基因的转基因细胞。
通过基因克隆实验,我们可以获得不同生物体的目标基因,并进行后续的研究和应用。
例如,通过将某种植物的耐旱基因克隆到其他作物中,可以提高作物的抗旱能力,增加农作物产量。
二、基因表达实验基因表达实验是将目标基因在宿主细胞中进行转录和翻译,产生具有特定功能的蛋白质的过程。
基因表达实验是研究基因功能和制备重组蛋白等领域的重要手段。
基因表达实验主要包括以下几个步骤:1. 选择合适的表达系统:根据需要表达的蛋白质的性质和规模,选择合适的表达系统。
常用的表达系统包括细菌、酵母、哺乳动物细胞等。
2. 构建表达载体:将目标基因插入到表达载体中,通常使用限制性内切酶和DNA连接酶进行连接,并通过测序确保插入正确。
3. 细胞转染:将构建好的表达载体导入到宿主细胞中。
不同表达系统有不同的转染方法,如细菌的化学转型、酵母的电转染等。
4. 表达和纯化:经过一定时间的培养,宿主细胞会表达目标基因,合成目标蛋白质。
可以通过蛋白质纯化技术,如亲和层析、凝胶电泳等手段获得纯度较高的目标蛋白质。
第八章酵母基因工程详解

二、酵母转化系统
酵母宿主系统 酵母载体系统 酵母系统标记基因 酵母转化方法 外源DNA在酵母宿主中的命运
1、酵母宿主系统
用作模式真核生物的酵母宿主菌 用作外源基因表达的酵母宿主菌 提高重组蛋白表达产率的突变宿主菌 抑制超糖基化作用的突变宿主菌 减少泛素依赖型蛋白降解作用的突变宿主菌
用作模式真核生物的酵母宿主菌
GAL10 Promoter
GAL80
GAL4
UAS
GAL1
GAL7 GAL10
A、 将GAL4的启动子换成GAL10的诱导型强启动子 B、半乳糖诱导GAL4高表达,不受GAL80产物抑制,激活GAL1等高效转录
3)pho4TS-PHO5启动子
低温诱导、磷酸盐抑制
A、PHO5启动子在培养基缺磷酸盐时启动转录
酿酒酵母(Saccharomyces cereviasiae):
*无性繁殖(芽殖或裂殖)、单细胞、真核生物 *繁殖方式与原核类似,易于操作 *基因表达调控机理与高等真核类似
用作外源基因表达的酵母宿主菌
酿酒酵母: 最成熟的真核细胞表达系统,表达水平低,产物过度糖基化
甲醇酵母:可以利用甲醇作唯一碳源,表达水平高,产物糖基化更合理
B、PHO4基因编码产物是PHO5启动子的正调控因子
C、pho4TS 温度敏感,35℃时失活,PHO5关闭
D、pho4TS-PHO5启动子通过降温(23℃)诱导表达
2、酿酒酵母分泌系统
酿酒酵母系统常用分泌信号肽来源: 性结合因子:MF-α 酸性磷酸酯酶:PHO5 蔗糖酶:SUC2 杀手毒素因子:KIL
碱金属离子介导的酵母菌完整细胞的转化
细胞:带壁的完整细胞 原理:通过碱金属离子(如Li+等)、PEG和热休克处理诱
第8章-酵母基因工程---基因工程原理与技术---刘志国-课件

酵母菌是比较成熟的真核生物表达系统。
作为宿主细胞的酵母需满足的基本要求
①安全无毒,没有致病性。 ②遗传背景清楚,容易进行遗传操作。 ③外源DNA容易导入宿主细胞,转化效率高。 ④培养条件简单,容易进行高密度发酵。 ⑤有较强的蛋白质分泌能力。 ⑥有类似高等真核生物的蛋白质翻译后的修饰加工能 力。
含有酵母菌染色体DNA同源序列的YIp质粒的构建
在大肠杆菌质粒上组装酵母菌染色体DNA特定序列和标 记基因,构建出来的质粒称为YIp。目的基因表达盒通常插 在染色体DNA特定序列中,这样目的基因就能高效整合入 酵母菌特定的染色体DNA区域。
酵母附加体质粒YEp:含有酿酒酵母2m质粒DNA复 制有关的序列,该载体在酵母细胞中稳定,拷贝数 可达60-100。转化效率高(b)。
REP1
A
IR
ori IR
同源重组
接合酵母属中的pSR1和pSB1,以及
克鲁维酵母属中的pKD1等均与2m质
B
粒类似。
FLP REP2
第一节 酵母基因工程表达体系 --------载体
酵母质粒载体既可以在大肠杆菌复制与扩增、又可以 在酵母系统中复制与扩增,故此类载体又称为穿梭载体( shuttle vector)。
由于巴斯德毕赤酵母没有合适的自主复制型载体,所以 外源基因序列一般整合入受体的染色体DNA上。其外源基因 的高效表达在很大程度上取决于整合拷贝数的多寡。目前已有 20余种具有经济价值的重组蛋白在该系统中获得成功表达。
多型汉逊酵母表达系统
多型汉逊酵母也是一种甲基营养菌。其自主复制序列 HARS已被克隆,并用于构建克隆表达载体, HARS质粒 能高频自发地整合在受体的染色体DNA上(可连续整合100多 个拷贝,因此重组多型汉逊酵母的构建也是采取整合的策略。
8基因表达,大肠杆菌基因工程

核糖体结合位点
核糖体结合位点的结构
大肠杆菌核糖体结合位点包括下列四个特征结构要素:
位于翻译起始密码子上游的6-8个核苷酸序列5’ UAAGGAGG 3’
,即Shine-Dalgarno(SD)序列,它通过识别大肠杆菌核糖体小 亚基中的16S rRNA 3’端区域3’ AUUCCUCC 5’并与之专一性结合, 将mRNA定位于核糖体上,从而启动翻译; 翻译起始密码子:以AUG;GUG或UUG作为翻译起始密码子; SD序列与翻译起始密码子之间的距离及碱基组成; 基因编码区5’ 端若干密码子的碱基序列。
C末端的丙氨酸交换下来,所形成的人胰岛素叔丁酯再用三氯乙酸脱
去叔丁酯基团,最终获得人胰岛素。该过程的总转化率为60%,工 艺路线耗时,分离纯化操作复杂,产品的价格不菲。
人胰岛素的生产方法
利用基因工程菌发酵生产人胰岛素
1982年,美国Ely LiLi公司首先使用重组大肠杆菌生产人胰岛素, 成为世界上第一个上市的基因工程药物;1987年,Novo公司又推出
A链和B链分别表达法
基因工程菌的构建战略: 化学合成A链 和B链的编码
Apr
tac
Met b-Gal Met A peptide Apr
tac
Met b-Gal Met B peptide
序列
ori
ori
M
M
M
M
N
C
N
C
重组人胰岛素的大肠杆菌工程菌的构建
A链和B链分别表达法
表达产物的后处理路线:
b-Gal
外源基因在原核细胞中的表达
现将真核基因在原核细胞中表达: 1、外源基因克隆在表达载体并导人宿主菌。
2、外源基因不能有间隔序列(内含子),因而必须用cDNA或全
大学《基因工程学》教学大纲
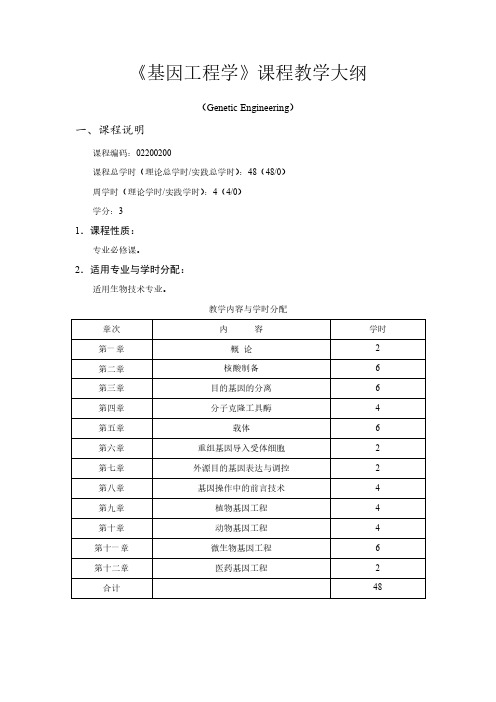
《基因工程学》课程教学大纲(Genetic Engineering)一、课程说明课程编码:02200200课程总学时(理论总学时/实践总学时):48(48/0)周学时(理论学时/实践学时):4(4/0)学分:31.课程性质:专业必修课。
2.适用专业与学时分配:适用生物技术专业。
教学内容与学时分配3.课程教学目的与要求:本课程的授课对象是生物技术专业的本科生。
课程简介:《基因工程》是生物技术专业的专业必修课程。
其以分子遗传学理论为基础,以分子生物学和微生物学的现代方法为手段而建立起来的一门技术学科。
基因工程兴起于20世纪70年代初,它的问世带动了生物技术的兴起和发展,是现代生物技术的核心内容。
基因工程课程的主要内容包括基因的分离、基因的克隆、基因的表达、植物基因工程、动物基因工程、药物基因工程和基因治疗等。
它是生命科学学院生物技术专业本科生的主干专业课程之一,它是生物工程(包括基因工程、细胞工程、酶工程、发酵工程)中最重要的课程,其它三大工程是建立在基因工程基础之上的,同时也为生物技术制药等后继学科奠定了重要的理论基础。
课程目标:设置本课程是为了让生物技术专业的学生理解和掌握基因工程的技术原理,通过本课程学习,掌握基因操作的工具酶,基因克隆常用载体,目的基因的分离与合成,重组体的构建,重组体向宿主细胞的导入,重组体克隆的筛选与鉴定以及克隆基因的表达,同时了解基因工程在生物学领域中的应用与发展前景。
对学生达到毕业要求贡献如下:1)了解基因工程学的历史、发展和前沿知识。
2)掌握基因工程学的基础理论、基本知识和基本技能;教学要求:学完基因工程学后,学生将具备以下能力:1)具有良好的自学能力;2)综合运用所掌握的基因工程学理论知识和技能、从事生物科学及其相关领域科学研究的能力。
4.本门课程与其它课程关系:先修课程为生物化学、微生物学、分子生物学、细胞学等,具备基础理论知识及实验能力是基因工程学课程的基础。
基因工程知识点全

第一章基因工程概述1.什么是基因工程,基因工程的基本流程基因工程Genetic engineering原称遗传工程;从狭义上讲,基因工程是指将一种或多种生物体供体的基因与载体在体外进行拼接重组,然后转入另一种生物体受体内,使之按照人们的意愿遗传并表达出新的性状;因此,供体、受体和载体称为基因工程的三大要素;1.分离目的基因2.限制酶切目的基因与载体3.目的基因和载体DNA在体外连接4.将重组DNA分子转入合适的宿主细胞,进行扩增培养5.选择、筛选含目的基因的克隆6.培养、观察目的基因的表达第二章基因工程的载体和工具酶1. 基因工程载体必须满足哪些基本条件➢具有对受体细胞的可转移性或亲和性;➢具有与特定受体细胞相适应的复制位点或整合位点;➢具有多种单一的核酸内切酶识别切割位点;➢具有合适的筛选标记;➢分子量小,拷贝数多;➢具有安全性;2. 质粒载体有什么特征,有哪些主要类型1、自主复制性2、可扩增性3、可转移性4、不相容性主要类型有1.克隆质粒2.测序质粒3.整合质粒4.穿梭质粒5.探针质粒6.表达质粒3. 质粒的构建1删除不必要的 DNA 区域,尽量缩小质粒的分子量,以提高外源 DNA 片段的装载量;一般来说,大于20Kb 的质粒很难导入受体细胞,而且极不稳定;2灭活某些质粒的编码基因,如促进质粒在细菌种间转移的 mob 基因,杜绝重组质粒扩散污染环境,保证 DNA 重组实验的安全,同时灭活那些对质粒复制产生负调控效应的基因,提高质粒的拷贝数3加入易于识别的选择标记基因,最好是双重或多重标记,便于检测含有重组质粒的受体细胞;4在选择性标记基因内引入具有多种限制性内切酶识别及切割位点的 DNA序列,即多克隆接头Polylinker,便于多种外源基因的重组,同时删除重复的酶切位点,使其单一化,以便环状质粒分子经酶处理后,只在一处断裂,保证外源基因的准确插入;5根据外源基因克隆的不同要求,分别加装特殊的基因表达调控元件;4. 什么是人工染色体载体将细菌接合因子、酵母或人类染色体上的复制区、分配区、稳定区与质粒组装在一起,即可构成染色体载体5. 什么是穿梭载体人工构建的、具有两种不同复制起点和选择标记、可以在两种不同的寄主细胞中存活和复制的载体;6.入-噬菌体载体及构建-DNA为线状双链DNA分子,长度为,在分子两端各有12个碱基的单链互补粘性末端;➢1缩短长度提高外源 DNA 片段的有效装载量删除重复的酶切位点➢引入单一的多酶切位点接头序列,增加外源DNA片段克隆的可操作性➢灭活某些与裂解周期有关基因;➢使λ-DNA载体只能在特殊的实验条件下感染裂解宿主细菌,以避免可能出现的污染现象的发生;➢加装选择标记,便于重组体的检测单链噬菌体DNA载体➢过定点诱变技术封闭重复的重要限制性酶切口;➢引入合适的选择性标记基因,如含有启动子、操作子和半乳糖苷酶氨基端编码序列lacZ’的乳糖操纵子片段lac、组氨酸操纵子片段his以及抗生素抗性基因等;➢将人工合成的多克隆位点接头片段插在 lacZ’标记基因内部,使得含有重组子的噬菌斑呈白色,而只含有载体 DNA 的混浊噬菌斑呈蓝色;➢4在多克隆位点接头片段的两侧区域改为统一的 DNA 测序引物序列,使得重组 DNA 分子的单链形式经分离纯化后,可直接进行测序反应;8. II类限制性内切核酸酶的特点限制性核酸内切酶 Restriction endonucleases是一类能在特异位点上催化双链DNA 分子的断裂,产生相应的限制性片段的核酸水解酶;➢识别位点的特异性:每种酶都有其特定的DNA识别位点,通常是由4、5或6核苷酸组成的特定序列靶序列;➢识别序列的对称性:靶序列通常具有双重旋转对称的结构,即双链的核苷酸顺序呈回文结构;➢切割位点的规范性:双链DNA被酶切后,分布在两条链上的切割位点旋转对称可形成粘性末端或平末端的DNA分子;同位酶:一部分酶识别相同的序列,但切点不同,这些酶称为同位酶;同裂酶:识别位点与切割位点均相同的不同来源的酶称为同裂酶同尾酶Isocandamers:识别位点不同,但切出的 DNA 片段具有相同的末端序列,这些酶称为同尾酶;9.甲基化酶Ⅱ类限制性内切酶有相应甲基化酶伙伴,甲基化酶的识别位点与限制性内切酶相同,并在识别序列内使某位碱基甲基化,从而封闭该酶切口;甲基化酶在封闭一个限制性内切酶切口的同时,却产生出另一种酶的切口➢甲基化酶可修饰限制性核酸内切酶识别序列,从而使DNA免受相应的限制性核酸内切酶的切割;➢甲基化酶的用途就是在必要时可以封闭某一限制性核酸内切酶的酶切位点;连接酶连接作用的特点:①DNA连接酶需要一条DNA链的3’末端有一个游离的羟基-OH,另一条DNA链的5’末端有一个磷酸基-P的情况下,只有在这种情况下,才能发挥连接DNA分子的作用;②只有当3’-OH和5’-P彼此相邻,并且各自位于与互补链上的互补碱基配对的两个脱氧核苷酸末端时,DNA连接酶才能将它们连接成磷酸二酯键;③DNA连接酶不能连接两条单链的DNA分子或环化的单链DNA分子,被连接的DNA链必须是双螺旋DNA分子的一部分;④DNA连接酶只能封闭双螺旋DNA上失去一个磷酸二酯键所出现的单链缺口nick,而不能封闭双链DNA的某一条链上失去一个或数个核苷酸所形成的单链裂口gap;⑤由于在羟基和磷酸基团之间形成磷酸二酯键是一种吸能反应,因此,DNA连接酶在进行连接反应时,还需要提供一种能源分子NAD+或ATP11.大肠杆菌 DNA聚合酶和Klenow大片段各有什么作用DNA聚合酶作用的特点:➢要有底物4种dNTP为前体催化合成DNA;➢接受模板指导;➢需要有引物3’羟基的存在;➢不能起始合成新的DNA链;➢催化dNTP加到生长中的DNA链3’-OH末端;➢催化DNA的合成方向是5’→3’;Klenow酶的基本性质:➢大肠杆菌DNA聚合酶I经胰蛋白酶或枯草杆菌蛋白酶部分水解生成的C末端604个氨基酸残基片段,即Klenow酶;分子量为76kDa;➢Klenow酶仍拥有5’→3’的DNA聚合酶活性和5’→3’的核酸外切酶活性,但失去了5’→3’的核酸外切酶活性;Klenow酶的基本用途:➢修复由限制性核酸内切酶造成的 3’凹端,使之成为平头末端;➢以含有同位素的脱氧核苷酸为底物,对DNA片段进行标记;➢用于催化 cDNA 第二链的合成;➢用于双脱氧末端终止法测定 DNA 的序列;聚合酶T4-DNA聚合酶酶的基本特性:➢有3’→5’的核酸外切酶活性和5’→3’的DNA聚合酶活性;➢在无dNTP时,可以从任何3’-OH端外切;➢在只有一种dNTP时,外切至互补核苷酸;➢在四种dNTP均存在时,聚合活性占主导地位;T4-DNA聚合酶的基本用途:切平由核酸内切酶产生的3’粘性末端13. 影响连接效率的因素有:➢温度最主要的因素离子浓度➢ATP的浓度 10μM - 1μM➢连接酶浓度平末端较粘性末端要求高➢反应时间通常连接过夜➢插入片段和载体片段的摩尔比➢DNA末端性质➢DNA片段的大小14.如何将不同DNA分子末端进行连接1.相同粘性末端的连接如果外源DNA与载体DNA均用相同的限制性内切酶切割,则不管是单酶酶解还是双酶联合酶解,两种DNA分子均含有相同的粘性末端,因此混合后能顺利的连接成一个重组DNA分子 2.平头末端的连接T4-DNA连接酶在ATP和高浓度酶的条件下,能连接具有完全碱基配对的平末端DNA分子,但平末端连接效率不高,基因操作不经常采用;3.不用粘性末端的连接3’端的粘性末端用T4-DNA聚合酶切平5’端的粘性末端用klenow酶补平,或者用S1核酸酶切平最后用T4-DNA连接酶进行平末端连接15. 碱性磷酸酶有什么作用1.该酶用于载体 DNA的5’末端除磷操作,以提高重组效率;2.用于外源DNA片段的5’端除磷,则可有效防止外源 DNA 片段之间的连接;16. 末端脱氧核苷酸转移酶有哪些作用➢给载体或目的DNA加上互补的同聚物尾;➢DNA片段3’末端的同位素标记;17. 2、细菌转化的步骤:∙感受态的形成;感受态时细胞表面出现各种蛋白质和酶类,负责转化因子的结合、切割及加工;感受态细胞能分泌一种小分子量的激活蛋白或感受因子,其功能是与细胞表面受体结合,诱导某些与感受态有关的特征性蛋白质如细菌溶素的合成,使细菌胞壁部分溶解,局部暴露出细胞膜上的 DNA 结合蛋白和核酸酶等;∙转化因子的结合;受体菌细胞膜上的DNA结合蛋白可与转化因子的双链DNA结构特异性结合,单链DNA或RNA双链RNA以及DNA/RNA杂合双链都不能结合在膜上;∙转化因子的吸收;双链 DNA 分子与结合蛋白作用后,激活邻近的核酸酶,一条链被降解,而另一条链则被吸收到受体菌中;∙整合复合物前体的形成;进入受体细胞的单链 DNA 与另一种游离的蛋白因子结合,形成整合复合物前体结构,它能有效地保护单链DNA免受各种胞内核酸酶的降解,并将其引导至受体菌染色体DNA处;∙转化因子单链DNA的整合;供体单链DNA片段通过同源重组,置换受体染色体DNA的同源区域,形成异源杂合双链 DNA结构;+诱导转化原理:①在0℃的Cacl2低渗溶液中,细菌细胞发生膨胀,同时Cacl2使细胞膜磷脂层形成液晶结构促使细胞外膜与内膜间隙中的部分核酸酶解离开来,诱导大肠杆菌形成感受态;②Ca2+能与加入的DNA分子结合,形成抗DNA酶DNase的羟基-磷酸钙复合物,并黏附在细菌细胞膜的外表面上;当42℃热刺激短暂处理细菌细胞时,细胞膜的液晶结构发生剧烈扰动,并随之出现许多间隙,为DNA分子提供了进入细胞的通道;③Mg2+对DNA分子有很大的稳定性作用,因此利用Mgcl2与Cacl2共同处理大肠杆菌细胞,可以提高DNA的转化效率;∙但该法要求条件高,对外界污染物极为敏感,通常很少采用;介导细菌的原生质体转化∙PEG是乙二醇的多聚物, 存在不同分子量的多聚体,它可改变各类细胞的膜结构, 使两细胞相互接触部位的膜脂双层中脂类分子发生疏散和重组,此时相互接触的两细胞的胞质沟通成为可能,从而造成细胞之间发生融合;20.电穿孔法是指在细胞上施加短暂、高压的电流脉冲,在质膜上形成纳米大小的微孔,DNA直接通过这些微孔或者作为微孔闭合时所伴随发生的膜组分重新分布通过质膜进入细胞质中,这种方法称为电穿孔法;P52 接合转化,入噬菌体感染未归纳21.转化率的影响因素.载体及重组DNA方面载体本身的性质:不同的载体转化同一株受体细胞,其转化率不同;载体的空间构象:与受体细胞亲和性较强的质粒载体转化率要高于亲和性较弱的质粒载体; 插入片段大小:对质粒载体而言,插入片段越大,转化效率越低;重组DNA分子的浓度和纯度受体细胞方面:受体细胞必须与载体相匹配转化操作的影响22.转化细胞的扩增转化细胞的扩增操作:指转化完成之后细胞的短时间培养;在实验时,扩增操作往往与转化操作偶联在一起,如:∙Ca2+诱导转化后的37℃培养一个小时∙原生质体转化后的再生过程∙λ噬菌体转染后的30℃培养等,均属扩增操作扩增操作的目的∙增殖转化细胞,使得有足够数量的转化细胞用于筛选程序;∙扩增和表达载体分子上的标记基因,便于筛选;∙表达外源基因,便于筛选和鉴定;23.抗药性筛选法这是利用载体DNA分子上的抗药性选择标记进行的筛选方法;抗药性筛选法的基本原理:抗药性筛选法可区分转化子与非转化子、重组子与非重组子将外源DNA片段插在EcoRI位点:∙非重组子呈 Apr、Tcr∙重组子呈 Apr、Tcr将外源DNA片段插在BamHI位点:∙非重组子呈 Apr、Tcr∙重组子呈 Apr、Tcs抗药性筛选法的基本操作:先将转化液涂布含有Ap的平板再将Ap平板上的转化子影印至含有Tc的平板上在Ap平板上生长,但在Tc平板上不长的转化子即为重组子 P56抗药性标记插入失活选择法∙经过上述抗药性筛选获得的大量转化子中既包括需要的重组子,也含有不需要的非重组子;为了进一步筛选出重组子,可利用质粒载体的双抗药性进行再次筛选;如果外源基因插入在载体的抗药性基因中间使得该抗药性基因失活,这种抗药性标记就会消失,从而筛选出阳性重组子;24. 什么是蓝白斑筛选法这种方法是根据组织化学的原理来筛选重组体;主要是在λ载体的非必要区插入一个带有大肠杆菌β—半乳糖苷酶的基因片段,携带有lac基因片段的λ载体转入lac的宿主菌后,在含有5—溴—4—氯—3—引哚—β—D—半乳糖苷X-gal平板上形成浅蓝色的噬菌斑;外源基因插人lac或lac基因部分被取代后,重组的噬菌体将丧失分解X-gal的能力,转入lac-宿主菌后,在含有5—溴—4—氯—3—引哚—β—D—半乳糖苷 X-gal平板上形成白色的噬菌斑,非重组的噬菌体则为蓝色噬菌斑;筛选法利用合适的引物,以从初选出来的阳性克隆中提出的质粒为模板进行PCR,通过对PCR产物的电泳分析,确定目的基因是否插入到载体中;由于在载体DNA分子中,外源DNA插入位点的两侧序列多数是已知的,可以设计合成相应的PCR引物,以待鉴定的转化子或重组子的DNA为模板进行PCR反应,反应产物经琼脂糖凝胶电泳,若出现特异性扩增DNA带,并且其分子量同预期的一致,则可确定含此重组DNA分子的重组子是期待的重组子;第三章基因工程的常规技术1. 探针有哪些类型探针标记有哪些方法类型:同源或部分同源探针cDNA探针人工合成的寡核苷酸探针标记方法:①5’端标记法②反转录标记法③缺刻前移标记法④ABC标记法4.什么是ABC荧光显色酶标记法ABC 标记法;∙A为Avidin生物素抗性蛋白,每个Avidin分子可结合3 - 4个生物素分子;∙B为Biotin生物素,每个Biotin分子可结合2个Avidin分子;∙C为Complex,首先将Biotin共价结合在探针分子上,荧光胺标记在Avidin上,两者形成复合物,即可将荧光胺标记在探针上,发出的荧光也能使普通胶片感光;如果将某一生色酶接在Avidin上,并辅以合适底物,则杂交反应还可直接以颜色反应检测,这一技术称为酶标技术5.亚克隆法∙亚克隆:是将克隆片段进一步片段化后再次进行的克隆;∙一般是将重组DNA分别用几种限制性核酸内切酶切割后,将所得各片段分别重组到载体上再转化宿主细胞,然后通过转化细胞的表型鉴定或鉴定,获得含有目的基因的重组子;此时,该重组分子中的无关DNA区域以被大量删除;6. 菌落嗜菌斑原位杂交的基本原理、流程∙该项技术是直接把菌落印迹转移到硝酸纤维素滤膜上,经溶菌和变性处理后使DNA 暴露出来并与滤膜原位结合再与特异性DNA探针杂交,筛选出含有插入序列菌落;∙操作步骤:∙①菌落生长∙②转移到NC膜上∙③DNA释放和变性∙变成单链DNA:∙ 10%SDS NaOH∙④中和 Tris-HCl pH∙⑤固定 80 ℃ 120’∙⑥杂交包括预杂交,加探针DNA杂交∙⑦放射自显影∙⑧对照比较,选出重组克隆7.鸟枪法∙鸟枪法:将某种生物体的全基因组或单一染色体切成大小适宜的 DNA 片段,分别连接到载体 DNA上,转化受体细胞,形成一套重组克隆,从中筛选出含有目的基因的期望重组子;鸟枪法制备目的基因的主要步骤∙①目的基因组DNA片段的制备超声波处理:片段长度均一,大小可控,平头末端;原核生物的基因长度大都在2Kb以内,真核生物的基因长度变化很大,最大的基因可达100Kb以上;全酶切:片段长度不均一,粘性末端便于连接,但有可能使目的基因断开,大小不可控;部分酶切:片段长度可控,含有粘性末端,目的基因完整;∙②DNA片段与载体连接如果转化子采用菌落原位杂交法或限制性酶切图谱法筛选,则选择多拷贝克隆载体;如果转化子采用基因产物功能检测法筛选,则选择表达型载体;∙③重组DNA分子导入受体细胞如果转化子采用菌落原位杂交法或限制性酶切图谱法筛选,则选择大肠杆菌作为受体细胞;如果转化子采用基因产物功能检测法筛选,则选择能使目的基因表达的受体细胞;∙④筛选含有目的基因的目的重组子菌落原位杂交法、基因产物功能检测法筛选模型的建立;∙⑤目的基因的定位利用鸟枪法获得的期望重组子只是含有目的基因的 DNA 片段,必须通过次级克隆或插入灭活,在已克隆的 DNA 片段上准确定位目的基因,然后对目的基因进行序列分析,搜寻其编码序列以及可能存在的表达调控序列;法酶促逆转录主要用于合成分子质量较大,转录产物mRNA易分离的目的基因;这种方法以目的基因的mRNA为模板,在逆转录酶的作用下合成互补的DNA,即cDNA,然后在DNA聚合酶的催化下合成双链cDNA片段,与适当的载体重组后转入受体菌扩增,获得目的基因的cDNA克隆; 的分离纯化绝大多数的真核生物mRNA在其3’端都存在一个多聚腺苷酸的尾巴,利用它可以迅速的将mRNA从细胞总的混合物中分离出来,将寡聚脱氧胸腺嘧啶共价交联在纤维素分子上,制成亲和层析柱,然后将细胞总的RNA混合物上层析柱分离,mRNA会挂在层析住上,后洗脱即可分离10. 简述PCR技术的基本原理,PCR反应体系的主要成分与主要程序是怎样的PCR技术的基本原理:类似于DNA的天然复制过程,其特异性依赖于与靶序列两端互补的寡核苷酸引物;过程:PCR由变性--退火--延伸三个基本反应步骤构成:①模板DNA的变性:模板DNA经加热至93℃左右一定时间后,使模板DNA双链或经PCR 扩增形成的双链DNA解离,使之成为单链,以便它与引物结合,为下轮反应作准备;②模板DNA与引物的退火复性:模板DNA经加热变性成单链后,温度降至55℃左右,引物与模板DNA单链的互补序列配对结合;③引物的延伸:DNA模板--引物结合物在TaqDNA聚合酶的作用下,以dNTP为反应原料,靶序列为模板,按碱基配对与半保留复制原理,合成一条新的与模板DNA 链互补的半保留复制链;重复循环变性--退火--延伸三过程,就可获得更多的“半保留复制链”,而且这种新链又可成为下次循环的模板;每完成一个循环需2~4分钟, 2~3小时就能将待扩目的基因扩增放大几百万倍;11. 什么是基因组文库其构建方法是怎样的是指将某种生物的全部基因组的遗传信息贮存在可以长期保存的稳定的重组体中,以备需要时能够随时应用它分离所需要的目的基因,这种保存基因遗传信息的材料,就称为基因文库又称DNA文库;基因组文库构建的一般步骤①载体的选择和制备;②高纯度、大分子量基因组 DNA 的提取;③基因组 DNA 的部分酶切与分级分离;④载体与DNA片段的连接;⑤转化或侵染宿主细胞;⑥筛选鉴定基因组及保存;12. 基因组DNA文库的质量标准除了尽可能高的完备性外,一个理想的基因组DNA文库应具备下列条件:∙重组克隆的总数不宜过大,以减轻筛选工作的压力∙载体的装载量最好大于基因的长度,避免基因被分隔克隆;∙克隆与克隆之间必须存在足够长度的重叠区域,以利于克隆排序;∙克隆片段易于从载体分子上完整卸下;∙重组克隆能稳定保存、扩增、筛选;基因文库的构建通常采用鸟枪法和cDNA法13.外源DNA片段的切割原则片段之间要有一定的重叠序列片段大小要均一文库构建的步骤∙细胞总RNA的提取和mRNA的分离∙第一链cDNA合成∙第二链cDNA合成∙双链cDNA的分级分离∙双链cDNA克隆进质粒或噬菌体载体并导入宿主中繁殖∙重组体的筛选与鉴定第四章基因在大肠杆菌、酵母的高效表达1. 启动子∙启动子:是DNA链上一段能与RNA聚合酶结合并能起始转录的序列,其大小在20~300个碱基,是控制基因转录的重要调控元件;在一定条件下mRNA的合成速率与启动子的强弱密切相关,而转录又在很大程度上影响基因的表达;∙启动子的特征:①序列特异性②方向性③位置特性④种属特异性2.启动子类型∙组成型启动子:是指在该类启动子控制下,结构基因的表达大体恒定在一定水平上,在不同组织、部位表达水平没有明显差异;∙组织特异启动子:又称器官特异性启动子;在这类启动子调控下,基因往往只在某些特定的器官或组织部位表达,并表现出发育调节的特性;∙诱导型启动子:是指在某些特定的物理或化学信号的刺激下,该种类型的启动子可以大幅度地提高基因的转录水平;目前已经分离了光诱导表达基因启动子、热诱导表达基因启动子、创伤诱导表达基因启动子、真菌诱导表达基因启动子和共生细菌诱导表达基因启动子等;3.终止子终止子:是位于结构基因下游的一段DNA序列,基因转录时,该序列被转录为mRNA的一部分,并形成特殊的二级结构,由此终止基因的转录;序列SD序列:mRNA中起始密码子上游8-13个核苷酸处有一段富含嘌呤核苷酸的顺序,它可以与30S亚基中的16S rRNA 3’端富含嘧啶的尾部互补,形成氢键结合,有助于mRNA的翻译从起始密码子处开始5.密码子不同生物对密码子的偏爱性1.生物体基因组中的碱基含量2.密码子与反密码子的相互作用的自由能3.细胞内tRNA的含量6. 密码子偏爱性对外源基因表达的影响∙由于原核生物和真核生物基因组中密码子的使用频率具有较大程大的差异性,因此外源基因尤其是高等哺乳动物基因在大肠杆菌中高效翻译的一个重要因素是密码子的正确选择;一般而言,有两种策略可以使外源基因上的密码子在大肠杆菌细胞中获得最佳表达:∙外源基因全合成∙同步表达相关tRNA编码基因7. 包涵体及其性质在某些生长条件下,大肠杆菌能积累某种特殊的生物大分子,它们致密地集聚在细胞内,或被膜包裹或形成无膜裸露结构,这种水不溶性的结构称为包涵体8. 包涵体的形成机理∙①折叠状态的蛋白质集聚作用;∙②非折叠状态的蛋白质集聚作用∙③蛋白折叠中间体的集聚作用;9. 包涵体的分离检测∙包涵体的分离主要包括菌体破碎、离心收集以及清洗三大操作步骤;10. 分泌型目的蛋白表达系统的构建∙包括大肠杆菌在内的绝大多数革兰氏阴性菌不能将蛋白质直接分泌到胞外,但有些革兰氏阴性菌能将细菌的抗菌蛋白细菌素分泌到培养基中,这一过程严格依赖于细菌素释放蛋白,它激活定位于内膜上的磷酸酯酶A,导致细菌内外膜的通透性增大∙因此,只要将细菌素释放蛋白编码基因克隆在一个合适的质粒上即可构建完全分泌型的受体细胞;此时,用另一种携带大肠杆菌信号肽编码序列和目的基因的表达质粒转化上述完全分泌型受体细胞,并使用相同性质的启动子介导目的基因的转录,则可实现目的蛋白从重组大肠杆菌中的完全分泌;11融合蛋白表达质粒的构建原则:∙受体细胞的结构基因能高效表达,且其表达产物可以通过亲和层析进行特异性简单纯化;。
分子生物学总结

分⼦⽣物学总结分⼦⽣物学总结第⼀章绪论⼀. DNA重组技术和基因⼯程技术.DNA重组技术⼜称基因⼯程,⽬的是将不同的DNA⽚段按照⼈们的设计定向连接起来,在特定的受体细胞中与载体同时复制并得到表达.产⽣影响受体细胞的新的遗传性状.基因⼯程技术还包括其他可能使⽣物细胞基因组结构得到改造的体系.第⼆章染⾊体与DNA⼀. DNA的⼀、⼆、三级结构特征.DNA⼀级结构特征1. 双链反向平⾏配对⽽成2. 脱氧核糖和磷酸交替连接,构成DNA⾻架,碱基排在内侧3. 内侧碱基通过氢键互补形成碱基对DNA⼆级结构特征绕DNA双螺旋表⾯上出现的螺旋沟,宽的沟称为⼤沟,窄沟称为⼩沟。
⼤沟,⼩沟都、是由于碱基对堆积和糖-磷酸⾻架扭转造成的。
DNA三级结构特征拓扑异构酶拓扑异构酶负超螺旋松弛DNA 正超螺旋溴已啶溴已啶⼆. 原核⽣物DNA具有哪些不同于真核⽣物DNA的特征.1. 结构简练2. 存在转录单元3. 有重叠基因三. DNA复制通常采取哪些⽅式.1. 线性DNA双链的复制.2. 环状DNA双链的复制分为θ型、滚环型和D-环型等.四. 原核⽣物DNA的复制特点.1. DNA双螺旋的解旋2. DNA复制的引发3. 冈崎⽚段与半不连续复制4. 复制的终⽌5. DNA聚合酶五. 细胞通过哪⼏种修复系统对DNA损伤进⾏修复?1. 错配修复2. 碱基切除修复3. 核苷酸切除修复4. DNA直接修复六. 什么是转座⼦?可分为哪些种类?转座⼦是存在与染⾊体DNA上可⾃主复制和位移的基本单位原核⽣物转座⼦的类型: 1. 插⼊序列 2. 复合转座⼦ 3. TnA家族第三章⽣物信息的传递(上)⼀. 什么是编码链?什么是模板链?与mRNA序列相同的那条DNA链称为编码链;将另⼀条根据碱基互补原则指导mRNA合成的DNA链称为模板链。
三. 简述σ因⼦的作⽤.σ因⼦的作⽤是负责模板链的选择和转录的起始,它是酶的别构效应物,使酶专⼀性识别模板上的启动⼦.四. 什么是Pribnow box?它的保守序列是什么?RNA聚合酶全酶与模板DNA结合后,⽤DNase I⽔解DNA,然后⽤酚抽提,沉淀纯化DNA后得到⼀个被RNA聚合酶保护的DNA⽚段,约有41-44个核苷酸对.在被保护区内有⼀个由5个核苷酸组成的共同序列,是RNA聚合酶的紧密结合点,称为Pribnow box. Pribnow区的保守序列是: TTGACA五. 简述原核⽣物和真核⽣物mRNA的区别.(⼀)原核⽣物mRNA的特征1、半衰期短2、多以多顺反⼦的形式存在3、5’ 端⽆“帽⼦”结构, 3’ 端没有或只有较短的polyA 结构。
【微生物工程】第八章_基因工程菌的培养

基因工程菌培养方式
连续培养 连续培养是将种子接入发酵反应器中,搅拌培养至菌体浓度达到一定程
度后,开动进料和出料蠕动泵,以一定稀释率进行不间断培养。 连续培养可以为微生物提供恒定的生活环境,控制其比生长速率,为研
究基因工程菌的发酵动力学、生理生化特性、环境因素对基因表达的影响等 创造了良好的条件。
但是由于基因工程菌的不稳定性,连续培养比较困难。为了解决这一问 题,人们将工程菌的生长阶段和基因表达阶段分开,进行两阶段连续培养。 在这样的系统中关键的控制参数是诱导水平、稀释率和细胞比生长速率。优 化这三个参数以保证在第一阶段培养时质粒稳定,菌体进入第二阶段后可获 得最高表达水平或最大产率。
基因工程菌的稳定性
基因工程菌的遗传不稳定性的的产生机制
受体细胞中的限制修饰系统对外源重组 DNA 分子的降解 外源基因的高效表达严重干扰受体细胞正常的生长代谢
能量、物质的匮乏和外源基因表达产物的毒性诱导受体细胞产生 应激反应:关闭合成途径,启动降解程序 重组质粒在受体细胞分裂时的不均匀分配 这是元件促进重组分子的缺失重排
提高质粒稳定性的方法
提高基因工程菌稳定性的策略
施加选择压力 根据载体上的抗药性标记,向培养系统中添加相应的抗生素 药物和食品生产时禁止使用抗生素 加入大量的抗生素会使生产成本增加 添加一些容易被水解失活的抗生素,只能维持一定时间 添加抗生素选择压力对质粒结构不稳定无能为力 载体上的营养缺陷型标记,向培养系统中添加相应的营养组份 培养基复杂,成本较高
r1质粒上的parb基因引入表达型载体中其表达产物可以选择性地杀死由于分配不均匀所产生的无质粒细胞基因工程菌的稳定性基因工程菌的稳定性提高基因工程菌稳定性的策略施加选择压力根据载体上的抗药性标记向培养系统中添加相应的抗生素药物和食品生产时禁止使用抗生素加入大量的抗生素会使生产成本增加添加一些容易被水解失活的抗生素只能维持一定时间添加抗生素选择压力对质粒结构不稳定无能为力载体上的营养缺陷型标记向培养系统中添加相应的营养组份培养基复杂成本较高提高质粒稳定性的方法提高质粒稳定性的方法提高基因工程菌稳定性的策略分阶段控制培养因外源基因表达造成质粒不稳定时可以考虑将发酵过程分阶段控制在生长阶段使外源基因处于阻遏状态避免因表达造成不稳定性问题的发生
第八章基因工程电子教案

导入外源DNA片段而获得新的表型的过程。
目的基因的筛选和鉴定
(screening/selection)
• 遗传学方法
– 插入灭活法(insertion inactivation):抗药性标志选择 – 标志补救 表达产物与营养缺陷互补
• 3′→5′外切酶活性 • 5′→3′外切酶活性
3′→5′外 切 酶 活 性
5′
3′
3′
5′
5′→3′外 切 酶 活 性
末端脱氧核 苷酰转移酶 (TDT)
载体 vector
• DNA ,能在宿主细胞中进行自我复制和 表达
• 克隆载体、表达载体 • 原核载体: 质粒(pBR322,pUC…)
Eco RⅠ Hind Ⅲ
氨苄青霉素 抗性基因
(ampr)
Pst Ⅰ
pBR322
Bam HⅠ
Sal Ⅰ
四环素 抗性基因
(terr)
Ava Ⅰ
O ri
Pvu Ⅱ
常用的克隆载体
• λ噬菌体(λphage) • 基因组分三个区域: 左侧区、中间区(非必需
区)、右侧区 • DNA,替换型载体,外源DNA: 9~23kb • 常用: EMBL 系列、 λgt 系列、charon系列 • 粘性质粒(cosmid): λDNA的 cos区+质粒,双链
环状DNA,克隆容量: 40~50kb • M13噬菌体 • 最大优点: 产生单链DNA
特点:
cos:cohesiveend site
RF DNA: replicational form DNA
优点:
表达载体(expressing vector)
基因工程-基因表达载体构建(2)

(4)、检测目的基因是否进入受体细胞可以用 DNA分子杂交 方法,用 DNA与RNA杂交 方法检测目的基因是否转录,用 免疫( 抗原抗体反应 )法检测目的基因是否表达。另外也 让虫子啃食棉叶,观察虫子的存活状态。 可进行个体水平检测。如 4、基因拼接成功的原因 DNA都是双螺旋结构,基本组成单位都是脱 ;
(3)将目的基因导入受体细胞:基因工程中常用的受体细胞 有大肠杆菌、枯草杆菌、土壤农杆菌、酵母菌和动植物细胞 受精卵 等。动物常把 细胞作为受体细胞。导入植物细胞的 方法有 等;农杆菌转化 农杆菌转化法、基因枪法、花粉管通道法 染色体DNA 法可以将目的基因导入细胞并把其整合到受体细胞的 显微注射法 上,导入动物细胞的方法有 ;如果运载体是质 CaCl2 处理,以增大细菌 细胞壁 粒,受体细胞是细菌,一般是将细菌用 的通透性,使含有目的基因的重组质粒进入受体细胞。目的 基因导入受体细胞后,就可以随着受体细胞的繁殖而复制, 由于 细菌繁殖的速度非常 ,在很短的时间内就能够获 快 得大量的目的基因。
途径 方法
供体细胞中的DNA 中直接分离基因 鸟枪法
供体细胞中的DNA ↓限制酶 许多DNA片段 ↓载入 运载体 ↓导入 受体细胞 ↓ 外源DNA扩增, 产生特定性状 ↓分离 目的基因
人工合成基因(真核细胞) 反转录法
目的基因mRNA ↓反转录 单链DNA ↓合成 双链DNA(即目的基因)
根据已知氨基酸 序列合成DNA
氧核苷酸且遵循碱基互不配对原则
转基因表达成功的原因是生物 共用一套遗传密码 。 基因工程的意义: 。 目的性强;克服远源杂交不亲和的障碍。
实例:利用大肠杆菌生产人的胰岛素简要过程:
胰岛素是治疗糖尿病的特效药,长期以来只能依靠从猪、牛等动物的胰腺 中提取,1000Kg胰腺只能提取40-50g的胰岛素,其产量之低和价格之高可 想而知。能否用大肠杆菌生产人的胰岛素?如果能,如何实现?
基因工程中的基因克隆与表达

基因工程中的基因克隆与表达基因工程是一门涉及分子生物学、遗传学、生物化学等多个学科的综合性科学。
其中,基因克隆和基因表达是基因工程研究的两个重要方面。
本文将就基因克隆和基因表达的原理、方法及应用进行探讨。
一、基因克隆1.原理基因克隆是指将目标基因从其天然基因组或其他来源中分离出来,并将其插入到另一个载体(如质粒)中,使其能够在宿主细胞内复制和表达。
基因克隆的原理是基于DNA序列特异性杂交的方法,利用限制性内切酶切割目标DNA和载体DNA,然后将它们黏合在一起,形成重组DNA。
通过转形或感染,使重组DNA 进入宿主细胞内,并复制和表达。
2.方法基因克隆的方法主要有限制性酶切与黏合(RE-Mediated Ligation)、PCR(聚合酶链反应)、TA克隆和基因文库等。
限制性酶切与黏合是一种常用的基因克隆方法。
该方法利用限制性内切酶切割DNA,然后通过T4 DNA连接酶黏合在一起。
这种方法操作简单、效率高,但存在限制内切酶的局限性,无法应用于不同酶切位点的DNA。
PCR是用于复制DNA片段的重要方法,也可以用于基因克隆。
PCR方法可以在不使用限制酶的情况下,从任何源提取DNA片段,扩增需要的基因段,并使用酶切和连接技术插入到载体中。
TA克隆是指用于从PCR产物中克隆DNA的一种方法。
该方法利用了Taq聚合酶不完全特异性合成3'-末端斜伸的性质,使产生的末端序列与T自带的A进行互补配对,从而使PCR产物能够被直接连接到TA克隆载体上。
基因文库是一种重要的基因克隆技术,可以将许多目标基因同时克隆入同一载体中。
基因文库分为cDNA文库和基因组文库。
通过荧光筛选或选择性培养,可以从文库中筛选出感兴趣的基因。
3.应用基因克隆技术广泛应用于基因工程、疫苗制备、药物研发、作物改良、动物遗传改良、环境污染治理等领域。
例如,利用基因克隆技术可以创造出超级细菌、工业用酶、新型药物、高产优质作物等。
二、基因表达1.原理基因表达是指基因通过转录和翻译的过程,将DNA序列转化为蛋白质的过程。
基因工程-第9章-外源基因的表达

故又称为TATA盒或—10区。启动子来源不同,
Pribnow盒的碱基顺序稍有变化。
-35区,位于转录起始位点上游35bp处, 故称-35区,一般由10个碱基组成。一般认为, -35区是RNA聚合酶σ亚基的识别与结合位点。 当σ亚基附着在-35区后,便带动RNA聚合酶 的核心酶(core enzyme,无σ亚基的RNA聚合 酶)沿DNA链向转录起始方向滑动至Pribnow盒, 并与之接触,而一旦它们相互结合之后,σ亚
转录终止子,目的是为了稳定载体系统。因
为上游强的tac启动子控制的转录必须由强终
止子抑制,才不至于干扰与载体本身稳定性 有关的基因 表达。
2.分泌型克隆表达载体pIN Ⅲ系统
这个载体系统是以pBR322为基础构建的。
它带有大肠杆菌中最强的启动子之一,即
Ipp(脂蛋白基因)启动子。ห้องสมุดไป่ตู้启动子的下游装
1. 启动子 启动子是DNA链上一段能与RNA聚合酶
结合并能起始mRNA合成的序列,它是基因表 达不可缺少的重要调控序列。没有启动子, 基因就不能转录。
原核生物启动子是由两段彼此分开且又 高度保守的核苷酸序列组成,对mRNA的合成 极为重要。
Pribnow盒,位于转录起始位点上游5~
10bp,一般由6~8个碱基组成,富含A和T,
点,即有一段富含A/T的区域和一段富含G/
C的区域,G/C富含区域又具有回文对称结 构,这段终止子转录后形成的RNA具有茎环
结构,并且具有与A/T富含区对应的一串U。
4.衰减子
衰减子(attenuator)是指在某些前导序列中 带有控制蛋白质合成速率的调节区。在原核 生物中,一条mRNA分子常常编码数种不同的 多肽链。这种多顺反子mRNA的头一条多肽链 合成的起始点,同RNA分子的5’-P末端间的距 离可达数百个核苷酸。这段位于编码区之前 的 不 转 译 的 mRNA 区 段 , 叫 做 前 导 序 列 (1eader)。此外,在mRNA的3'-OH末端,以及 在多顺反子mRNA中含有的长达数百个碱基的 顺反子间序列(intercistranic-sequence),即间隔 序列 (spacer),也发现有不转译的序列。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
Transcript folded to form termination hairpin
终止子
强化转录终止的必要性
外源基因在强启动子的控制下表达,容易发生转录过头现象,即RNA聚合酶滑 过终止子结构继续转录质粒上邻近的列,形成长短不一的mRNA混合物 过长转录物的产生在很大程度上会影响外源基因的表达,其原因如下: 转录产物越长,RNA聚合酶转录一分子mRNA所需的时间就相应增加,外源基因 本身的转录效率下降; 如果外源基因下游紧接有载体上的其它重要基因或DNA功能区域,如选择性标 记基因和复制子结构等,则RNA聚合酶在此处的转录可能干扰质粒的复制及其它生 物功能,甚至导致重组质粒的不稳定性; 过长的mRNA往往会产生大量无用的蛋白质,增加工程菌无谓的能量消耗; 更为严重的是,过长的转录物往往不能形成理想的二级结构,从而大大降低外源 基因编码产物的翻译效率
E. coli
---------TTGACA--------TATAAT-----35 box -10 Pribnow box GENE
Animals
-----Various signal-------TATAAAT----25 box GENE
Commonly used promoters
The lac promoter:
The tac/trc promoter:
- A hybrid between the trp and lac promoters - Stronger than either and still induced by IPTG
启动子
启动子的可控性
乳糖启动子Plac的可控性:
野生型的Plac与其控制区Olac 偶联在一起,在没有诱导物 存在时,整个操纵子处于基 底水平表达;诱导物可以使 启动子Plac介导的转录大幅 提高 P O 高效转录 P 乳糖 异丙基--D-硫 代半乳糖苷(IPTG) 诱导 O 基底水平转录 阻遏蛋白
SD序列: mRNA中用于结合原核生物核糖体的序列。SD 序列在细菌mRNA起始密码子AUG上游10个碱基左右处, 有一段富含嘌呤的碱基序列,能与细菌16SrRNA3’端识别, 帮助从起始AUG处开始翻译。 启动子:DNA分子上能与RNA聚合酶结合并形成转录起始 复合体的区域,在许多情况下,还包括促进这一过程 的调节蛋白的结合位点。 终止子:常指转录终止子。①转录过程产生RNA的一段可 终止转录的茎-环结构序列;②位于模板基因下游该结 构所对应的DNA序列。在大肠杆菌中有依赖于ρ或不依 赖于ρ的两类终止子。 核糖体结合位点:核糖体识别并结合信使核糖核酸 (mRNA)的位点。原核生物信使核糖核酸起始密码子的 上游含有核糖体识别和结合序列;真核生物信使核糖 核酸的5′帽子结构对核糖体的识别起一定作用。
第八章 基因的表达
第一节 基因表达研究的方法 第一节 影响外源基因表达的因素
第一节 基因表达研究的方法
大肠杆菌作为基因工程宿主菌的优越性
大肠杆菌是迄今为止研究得最详尽的原核生物; 结构简单,生理生化和遗传背景知识,尤其是其基因 表达调控机制有了清楚的了解; 已被发展成为一种安全的基因工程实验体系,拥有各 种宿主菌和载体; 培养简单,繁殖迅速,培养代谢易于控制,易于进行 工业化批量生产; 实验已经证明,许多克隆的真核基因,如生长激素基 因和胰岛素基因等,都可在大肠杆菌中实现高效表达。
1. The Promoter
The promoter is the critical component of gene expression DNA sequence recognized by the RNA polymerase The bound polymerase initiates transcription downstream the DNA toward the gene There are strong and weak promoters
Plac
启动子
启动子的可控性
色氨酸启动子Ptrp的可控性:
色氨酸启动子Ptrp受色氨酸-阻遏 蛋白复合物的阻遏,转录呈基底 状态。在培养系统中去除色氨酸 或者加入3-吲哚丙烯酸(IAA), 便可有效地解除阻遏抑制作用。 在正常的细菌培养体系中,除去 色氨酸是困难的,因此基因工程 中往往添加IAA诱导Ptrp介导的目 基因的表达 Ptrp Otrp Ptrp Otrp 高效转录 阻遏蛋白 色氨酸 基底水平转录
Base pairing between a mRNA’s Shine Delgarno sequence and the 16s rRNA permit the ribosome to select the proper initiation codon The distance between SD sequence and the initiation codon of the gene is important to translation.
Ptrp 除去色氨酸
Otrp 或加3-吲哚丙烯酸 (IAA) 高效转录
启动子
启动子的可控性
阻遏作用
噬菌体启动子PLL的可控性:
目的基因 噬菌体启动子PLL受CI阻遏蛋白阻 Ptrp 遏,很难直接诱导控制。在基因 工程中常使用温度敏感型的cI突 变基因cI857控制PL。 cI857阻遏蛋 在42℃时失活脱落,PLL便可介导 目的基因的表达。但在大型细菌 培养罐中迅速升温非常困难,因 此常使用一个双质粒控制系统, 用色氨酸间接控制目的基因表达 Ptrp A 色氨酸 PLL B 表达 A cI857 PLL B
2. Terminator
Stops RNA synthesis at specific points along the DNA template The enzyme stops polymerizing nucleotides into RNA, releases the RNA chain and leaves the DNA to eventually reinitiate at another promoter A terminator is usually a nucleotide sequence that can base pair itself to form a stem loop structure. The transcription will stop at this point.
The three most important signals for gene expression in E.coli
Ribosome Promoter binding site
Terminator
Gene
DNA RNA transcript
Point at which ribosome binds to mRNA
Upstream sequence around position -35 also have a region of sequence similarity, TCTTGACAT which is most evident in efficient promoters
Typical promoter sequences for E.coli and animal genes
ori Aprr pCP1 Tcrr
筛选Aprr、Tcss的转化子
3. Ribosome binding site (RBS)
RBS can vary in E.coli
A consensus sequence (Shine-Delgarno sequence) is centered ~10 nucleotides upstream from the start codon Most sites contain the minimal sequence UAAGGAGG
The promoter : marks the point at which transcription of the gene should start The terminator : marks the point at the end of the gene where transcription should stop The ribosome binding site (RBS): a short nucleotide sequence recognized by the ribosome as the point at which it should attach to the RNA molecule
启动子与RNA聚合酶 聚合酶 启动子与
RNA聚合酶同启动子结合的区域称为启动子区。将各 种原核基因同RNA聚合酶全酶结合后,用DNase I水解 DNA,最后得到与RAN聚合酶结合而未被水解的DNA片 段,这些片段有一个由5个核苷酸(TATAA)组成的共同 序列,以其发现者的名字命名为Pribnow框 (Pribnowbox),这个框的中央位于起点上游10bp处, 所以又称—10序列(—10 sequence),后来在—35 bp 处又找到另一个共同序列(TTGACA)。 Hogness等在真核基因中又发现了类似Pribnow框的共 同序列,即位于—25~—30 bp处的TATAAAAG,也称 TATA框(TATAbox)。
启动子
启动子的可控性
乳糖启动子Plac的可控性:
野生型的Plac上游附近拥有 代谢激活因子(CAP)结合 区,cAMP激活CAP,CAP 结合启动子控制区,进而促 进Plac介导的转录。葡萄糖 代谢使cAMP减少,也能阻 遏Plac介导的转录。因此, 基因工程中使用的乳糖启动 子均为抗葡萄糖代谢阻遏的 突变型,即Plac UV5 Plac UV5 O Plac O 高效转录 葡萄糖代谢 CAP O cAMP浓度降低 基底水平转录 cAMP 高效转录
Main Features of E.coli Promoter