植物叶绿体中果糖1,6-二磷酸醛缩酶(Fructose 1,6 bisphosphate aldolase,FDA)试剂盒使用说明
果糖-1,6-二磷酸醛缩酶(FBA)检测

迪信泰检测平台
果糖-1,6-二磷酸醛缩酶(FBA)检测
果糖1,6-二磷酸醛缩酶(Fructose 1,6 bisphosphate aldolase, FBA)是糖酵解、糖异生、磷酸戊糖途径及光合作用中参与卡尔文循环的重要酶,催化果糖1,6-二
磷酸可逆的裂解为磷酸二羟丙酮和3-磷酸甘油醛,广泛存在于动植物及微生物体内,在各种逆境胁迫下表现不同的响应,能与细胞骨架结合,参与微管的聚合,细胞的内吞和膜泡运输,同时还参与了病原菌的侵染过程。
迪信泰检测平台采用生化法,结合相应的试剂盒,可实现高效、精准的果糖-1,6-
二磷酸醛缩酶的活性变化。
此外,我们还提供其他光合作用类的检测服务,以满足您的不同需求。
生化法测定果糖-1,6-二磷酸醛缩酶样本要求:
1. 请确保样本量大于0.2g或者0.2mL。
周期:2~3周。
项目结束后迪信泰检测平台将会提供详细中英文双语技术报告,报告包括:
1. 实验步骤(中英文)。
2. 相关参数(中英文)。
3. 图片。
4. 原始数据。
5. 果糖-1,6-二磷酸醛缩酶活性信息。
迪信泰检测平台可根据需求定制其他物质测定方案,具体可免费咨询技术支持。
叶绿体中Calvin循环多酶复合体

叶绿体中Calvin循环多酶复合体可溶性酶组合成多酶复合体的优势及其与类囊体膜的结合对Calvin循环多酶复合体的研究进展作了介绍.关键词:Calvin循环;多酶复合体;类囊体膜以往的研究认为,细胞及其细胞器内充满了游离分子,精确的化学反应是通过这些游离分子的非目的性碰撞而发生的,但现代科学家们对这个观点提出了质疑.目前,根据蛋白质浓度,蛋白质在胞液中的溶解能力,酶一酶之间的相互作用提出:代谢途径的酶在体内应该以复合体的形式存在,并且这种复合体与亚细胞结构和生物膜相结合,为”底物穿梭”提供更加便利的通道,继而提高体内蛋白拥挤的环境中的代谢效率(Srere1987).尽管可溶性酶的组合在蛋白质拥挤的环境中有显着的生理作用,但是用生物化学和分子生物学的技术证明这种观点是非常困难的,因此这种复合体的形成及其代谢重要性遂成为争论的主题.一些动物和微生物细胞中的多酶复合体己有广泛研究(Srere1980;Ovadi1988;Keleti等1989),但关于植物代谢途径中酶的组合信息则很有限(Hrazdina和Jensen1992),且大多数集中于电子传递链组分的研究.植物生物化学研究的目的之一就是提高植物体中代谢的效率,其中也包括作物的光合作用,因此了解体内这种多分子组合及其对代谢途径的调控就非常重要.本文对叶绿体基质中可溶性酶之间组合成多酶复合体以及可溶性酶与类囊体膜结合的研究进展作介绍.1Calvin循环.Calvin循环利用光合作用原初反应和氧释放,光合磷酸化以及电子传递的反应产物ATP和NADPH,将活跃的化学能转换为贮存在糖类中的稳定的化学能,以供给生命活动的需要,即碳同化.这个循环在叶绿体基质中进行,需要11种酶来催化13个连续的反应,分为3个阶段,即羧化阶段,还原阶段和更新阶段,具体的反应过I程如图l..长期以来一直认为Calvin循环中的各个酶都图lCalvin循环【根据Raines(2003)略加修改】A:l,5一二磷酸核酮糖羧化酶,加氧酶(Rubisco);B:3一磷酸甘油酸激酶(PGK);C:3一磷酸甘油醛脱氢酶(GAPDH);D:丙糖磷酸异构酶(TPI);E:醛缩酶(aldolase):F:果糖.1,6.二磷酸磷酸酶(FBPase);G:转酮酶(transketolase);H:景天庚酮糖一l,7二磷酸酶(SBPase);I:核酮糖.5一磷酸差向异构酶(Ru.5一Pepimerase);J:核糖磷酸异构酶(RPI);K:磷酸核酮糖激酶(RPK).3一PGA:3一磷酸甘油酸:1,3一PGA:1,3-二磷酸甘油酸:G.3一P:3一磷酸甘油醛:DHAp:二羟丙酮磷酸;Fru—l,6一P:果糖一l,6.二磷酸;Fru一6-P:果糖.6.磷酸;E.4一P:赤藓糖一4.磷酸;Sed—l,7-P:景天庚酮糖.1.7.二磷酸;Sed一7一P:景天庚酮糖一7一磷酸;R一5一P:核糖一5一磷酸;x. 5一P:木酮糖一5一磷酸;Ru一5一P:核酮糖.5一磷酸;Ru.1.5.P:核酮糖一l,5一二磷酸.收稿2007—12—07修定2008—04—10资助南京农业大学科技创新基金(Y200523.通讯作者(E—mail:************.cn;Tel:025.84395423,.植物生理学通讯第44卷第3期,2008年6月393是以可溶性状态游离存在于高等植物叶绿体中的,直到上个世纪60年代,Calvin循环中的酶相互结合形成多酶复合体才被发现(vanNoort和Wildman1964;Mendiola和Akazawa1964).目前,许多实验室已经从叶绿体中分离纯化得~lJCalvin循环一些酶组成的多酶复合体.2多酶复合体的分离与特性MUller(1972)认为一些CO,固定酶可能是以一种易解离的复合体形式存在的,但是一般可溶性酶之间的相互作用是弱而短暂的,因此它们的多分子组合的研究比较难,尽管有人曾用不同的手段观察到了Calvin循环酶之间的多酶组合.Sainis和Harris(1986,1987)根据蔗糖密度梯度离心分离得到的豌豆和菠菜叶绿体基质提取物中,1,5一二磷酸核酮糖羧化酶/加氧酶(Rubisco,EC4.1.1.39)组分可以核糖一5一磷酸(R一5一P)为底物表现出依赖ATP的羧化活性,这表明核糖磷酸异构酶(RPI,EC5.3.1.6),磷酸核酮糖激酶(RPK,EC2.7.1.19)与Rubisco可以作为一个整体而纯化出来,后又发现几乎所有RPK都可与羧化酶结合.Gontero等(1988)用离子交换层析,分子筛层析和羟基磷灰石柱层析方法从菠菜中纯化出一个由5个酶组成的复合体,这些酶是卡尔文循环过程中的连续酶,包括RPI,RPK,Rubisco,3一磷酸甘油酸激酶(PGK,EC2.7.2.3)和3一磷酸甘油醛脱氢酶(GAPDH,EC1.2.1.13),这个复合体的分子量为540kDa.由这5个酶组成的复合体催化的反应过程如下:核糖一5一磷酸垦核酮糖一5一磷酸垦坚核酮糖一1,5一二磷酸生!!.3一磷酸甘油酸1,3一二磷酸甘油酸GAPDH3一磷酸甘油醛+NAD++HPO Babadzhanova等(2002)在棉花叶片中也分离得到包括这5个酶的多酶复合体,此种复合体分为游离的和类囊体膜结合的两种形式,分子量分别为(520~20)kDa和(640~25)kDa.2006年他们(Babadzhanova等2006)又在生长15~25d(6~8真叶阶段,花芽形成期间,开花期间)的棉花叶片中分离出分子量分别为(240~lO)kDa和(520~20) kDa的游离多酶复合体,并对发育过程中的RPI, RPK和Rubisco的活性变化进行了比较研究,结果表明多酶复合体中的RPI和RPK与单一酶相比活性变化不显着,Rubisco活性变化显着,生殖器官形成阶段复合体酶活性和光合速率最高.Rault等(1993)研究由这5个酶组成的复合体的结构和功能特性的结果表明,此种复合体中每一酶组分的四级结构都不同于游离的单一酶的四级结构,其动力学研究(Gontero等1993)结果表明,复合体中的酶有较高的和较低的rm.Ricard等(1994)提出,不同的酶组合成多酶复合体后其催化特性发生变化,这种变化与不同活化位点之间的反应中间产物通道无关,而是多酶复合体中酶彼此之间信息传递的结果(RiCard等1994; Gontero等1994).迄今研究较多的是由GAPDH和RPK组成的多酶复合体,这2个酶在Calvin循环过程中催化不连续的反应,但也可以结合成为复合体而存在. Nicholson等(1987)在斜生栅列藻(Scenedesrnus obliquus)中分离得到一个稳定的含有GAPDH和RPK的复合体,分子量为560kDa,这2个酶呈结合状态,彼此之间的动力学特性相互影响.在衣藻(Chlamydomonsreinhardtii)的叶绿体中也分离和纯化出一个含有RPK和GAPDH的双酶复合体(Lebreton等1997;Avilan等1997),热力学统计的结果表明其蛋白之间有信息和能量传递.复合体的稳定或不稳定是通过构象变化来反映的,而这种构象变化则是通过蛋白质一蛋白质的相互作用表现出来,最终各组分之间产生互相结合的指纹.构象变化的热力学研究表明GAPDH和RPK即使在分离之后仍然携带有这些指纹,最终引起一些辅助因子的动力学特性发生变化(Lebreton等1997;Avilan等1997;Lebreton和Gontero1999; Graciet等2002).Mouche等(2002)采用多重技术相结合的方法研究衣藻(Chlamydomonasreinhardtii)中GAPDH和RPK组成的多酶复合体的结果表明, 二聚体的RPK在与四聚体的GAPDH结合和分离的循环过程中的活性有显着变化;低温电子显微镜观察表明分离的和模型化的RPK二聚体和整个复合体的三维结构的对应体之间在结构上有显着变化.RPK与GAPDH之间相互结合,并可以利用原初反应和氧释放,光合磷酸化和电子传递反应中产生的ATP和NADPH,此种双酶复合体中2个酶之间相互协同调控,定义为一个”控制单位”——Calvin循环是光,pH和代谢物调控的一个起始点.因此认为,RPK与GAPDH之间的蛋白质一394植物生理学通讯第44卷第3期,2008年6月蛋白质相互作用可更好地调控Calvin循环(Lebreton 和Gontero1999).近2年来还发现,GAPDH和RPK组成的多酶复合体还可与CP12共同组成一个多酶复合体, CP12是一个核基因编码的分子量为8.5kDa的叶绿体蛋白,在超分子复合体装配中起蛋白连接子的作用(Graciet等2003),GAPDH,RPK和CPI2的组装模型如图2所示,首先GAPDH与CP12相互结合,解离常数接近0.44nmo1.L~,这种结合可能使GAPDH构象发生改变,而后GAPDH/CP12复合体与RPK相互结合,其解离常数为60nmo1.L-, 最终形成半个复合体,定义为一个单位.这个单位以二聚体形式最终形成包括2个二聚体的RPK,2个四聚体的GAPDH,可能是2个单体CP12的复合体.Graciet等(2004)曾对GAPDH/CP12/RPK组成的复合体的动力学,调控特性及其在CO同化中的作用作了详细综述.Marri等(2005)也曾对拟南芥进行体外重组得到GAPDH/cP12/RPK相互结合的复合体,其分子量为640kDa.图2衣藻(Creinhardtii)的GAPDH,RPl(/CP12复合体组装模型【根据Graciet等(2003)略加修改】除了由5个酶和2个酶组成的多酶复合体之外,人们还发现了由其他不同酶组分组成的Calvin循环多酶复合体.Persson和Johansson (1989)用液体两相系统的逆流分配探测6个Calvin 循环酶的共分配现象的结果表明这些酶之间存在蛋白质.蛋白质复合体趋势,这些酶包括Rubisco, PGK,GAPDH,丙糖磷酸异构酶(TPI,EC5.3.1.1),醛缩酶,果糖.1,6.二磷酸磷酸酶(FBPase,EC3.1.3.11).Siiss等(1993)分离出一个含有RPI,RPK,Rubisco,GAPDH,景天庚酮糖.1,7一二磷酸酶(SBPase,EC3.1.3.37),铁氧还蛋白NADP还原酶(FNR,EC1.18.1.1)的多酶复合体,其分子量为900kDa.这一复合体在低离子强度溶液中比较稳定,Calvin循环酶形成催化链,以5.磷酸核糖为底物,经过多步连续酶促反应合成3.磷酸甘油酸,这种”底物穿梭”可大幅度的增加催化效率.Jebanathirajah和Coleman(1998)用快速蛋白液相色谱(fastproteinliquidchromatography, FPLC),从烟草叶绿体中分离得到碳酸酐酶(CA, EC4.2.1.1)与Rubisco,RPK,RPI相结合的多酶复合体,其分子量高于600kDa,CA可能与Rubisco 结合在卡尔文循环多酶复合体的外侧,RPK位于复合体中心,并与类囊体膜结合.总之,这些体外研究的结果表明,大部分参与Calvin循环的酶均可以结合成为多酶复合体,但不同复合体之间的酶组分则不完全相同,酶组分之间没有确定的化学计量比(表1).Rault等(1993)采用免疫化学和光密度分析相结合的方法研究由RPI,RPK,Rubisco,PGK和GAPDH组成的多酶复合体中酶组分的化学计量比时发现,多酶复合体含有RPK的2个亚基,RPI的2个亚基,Rubisco的2个大亚基(L)和4个小亚基(S), PGK的1个亚基,GAPDH的2个不同的亚基(0’亚基和B亚基).Hosur等(1分析多.993)Rubisco/RPK酶复合体的结晶和结构时,发现Rubisco是正常的S.形式,但对其中一些晶体进行X.ray衍射的结果表明,晶体Rubisco除正常LS形式之外还有其他的形式,如上面所说的LS韵形式,这可能是多酶复合体中其他酶影响造成的.根据不同实验室的结果可以认为,其原因可能是采用的提取和纯化程序不同造成的,也可能是多分子之间结合的松紧程度不同和动态变化造成的.体内由基质酶和膜结合的酶组成的蛋白复合体可能是蛋白网络排列的,而不是按单个酶组分”无秩序的”随机分布排列的,因此溶解于水介质中时的多酶复合体会趋向解离,各组分分开,只有非常稳定的酶复合体在提取之后才有可能在非生理的液相环境中仍然保持结合状态,分离出来可用于特征研究,否则就难以确定多酶复合体中酶组分的化学计量比.另外,分离Calvin循环中酶复合体时也不能排除实验操作过程中由于人为因素而引起部分游离植物生理学通讯第44卷第3期,2008年6月395 酶自由组合成多酶复合体,应该采用灵敏度高的检测方法证明新鲜细胞器提取物中的酶是聚合成复合体而不是人为因素造成的.Stiss等(1993)用限制性蛋白酶水解和免疫印迹相结合的方法证实了这一点.这个方法是建立在Lys和Arg残基通常参与寡聚蛋白蛋白交界面形成的基础之上的.如果能够切割这2种氨基酸的肽键进而使蛋白组分分开,则所有的样品都可在同一介质中进行蛋白酶水解,反过来,特定酶需要具有同一构型;与单一酶相比,多肽的差异应该是由蛋白质.蛋白质之间的相互作用引起的.单独的酶可能更容易受到特异性蛋白酶的影响和裂解,而如果这些组分结合到多酶系统中就不易受蛋白酶水解的影响. 经证明,专一的裂解精氨酸和赖氨酸残基的胰蛋白酶是这~实验中的常用品,有人用胰蛋白酶已成功证明IK和GAPDH在新鲜准备的基质提取物中是相互结合的.此外,在不同离子强度下Calvin循环酶复合体的解离也可采用这种技术加以证明(Stiss等1993).从这些结果中还可以看出,基质提取物的任何处理都可能引起Calvin循环多酶复合体的解离,这就可以解释为什么从同样的提取物中分离的Calvin循环多酶复合体有组分的差异和分子量高低之分.如同Calvin循环一样,其他代谢途径的连续酶也可以在叶绿体中结合成为多酶复合体,这样所有代谢反应才可以在一个协调的环境中进行,光反应也才可以更有效地利用能量,产生还原力,有关其他代谢途径中酶组合的信息尚少,还有待继续研究.3可溶性酶组合成多酶复合体的优势及其与类囊体膜的结合植物体内的一些代谢途径可以共享同样的代谢中间产物,并同时在特定的时间内于拥挤的环境中进行,因此将酶之间代谢途径的细微区域化与底物通道连接起来研究是非常必要的,这就可以将这些代谢途径的共同底物和辅助因子之间的竞争降到最低,代谢过程得以有效进行.SnSS等(1993)提出,多酶组合能够使CO,同化所需组分和辅助因子(A TP和NADPH)更加便利的运输到催化位点,还可以防止其他代谢途径的干扰.Rault等(1993)研究RPI,RPK,RubiSCO,PGK和GAPDH构成的复合体的动力学结果表明, Rubisco以复合体形式存在时,其催化效率提高,这可能是每一活化位点的.增加和下降造成的.Sainis和Jawali(1994)在体外观察到,由RPI,RPK,Rubisco组成的Calvin循环多酶复合体可以表现出以R.5.P为底物和依赖ATP的CO,固定活性,可为中间代谢物提供通道,致使Rubisco的催化反应更加便利.PGK和GAPDH在体外的相互结合表明二者之间可以相互作用,并可为l,3.二磷酸甘油酸的穿梭提供通道(Wang等1996).Brenda(2004)认为,酶结合成复合体后的最大优势就是可使一些生物合成的代谢中间产物更加便利的运输到催化位点,而不是在细胞内聚成团,这种代谢通道在提高和调控细胞的生物化396植物生理学通讯第44卷第3期,2008年6月学反应中发挥作用.一般,多酶复合体可以为“底物穿梭”提供更加便利的通道,从而提高酶催化效率,同时还可增加酶结构的稳定性,其在体内蛋白质拥挤的环境中的优势更明显.除了可溶性酶之间可以相互作用结合成复合体以外,可溶性酶还可与类囊体膜结合.已有的研究表明,Rubisco,FBPase,RPK,GAPDH,PGK和RPI可结合在类囊体膜上(Mori等1984; Hermoso等1989;Sainis和Melzer2005),I-I+一ATP 合酶可以为Rubisco提供膜结合位点(Sainis和Melzer2005).但实验过程中由于一些人工操作所引起的酶吸附在类囊体膜上的假象不能被排除,这一问题己可以通过低温固定叶切片和免疫电镜加以解决.早期的免疫化学研究表明,Rubisco分子随机分布在叶绿体基质中(Shaw和Henwood 1985;V aughn1987),其他的卡尔文循环酶也是均匀分布的,但有人用免疫电镜研究的结果对此种假设提出了质疑,发现卡尔文循环中的酶,如:RPK,GAPDH和SBPase都结合在类囊体膜上(Stiss等1993).此外,还有人发现微藻中的RPK 也结合在类囊体膜上(Mangeney等1987),更有人发现FBPase和硫氧还蛋白也结合在类囊体膜上(Hermoso等1992).在衣藻的叶绿体中用免疫电镜可以观察到Calvin循环酶,如RPI,GAPDH, FBPase,醛缩酶和PGK在体内均与类囊体膜结合(SUss等1995).豌豆叶片的免疫定位观察表明, 60%的GAPDH和54%的PGK结合于类囊体膜上(Wang等1996).而Anderson等(1996)对豌豆叶片进行免疫定位后发现大部分的PGK,SBPase,一半的GAPDH,RPK,小部分的TPI,醛缩酶,CA都是与类囊体膜结合的.此外Teige等(1998)报道Calvin循环中的一些次要酶类型,如:核酮糖一5一磷酸差向异构酶,转酮酶也都结合在类囊体膜上. FNR可与包括GAPDH在内的Calvin循环多酶复合体一同分离得~U(SUss等1993),它也是与类囊体膜结合的电子传递链的组分之一,因此推测PSI和Calvin循环之间有密切关系.GAPDH是卡尔文循环中唯一依赖NAPDH的酶,但在基质提取物的免疫亲和层析中用抗FNR和抗GAPDH的抗体的结果表明二者是相互结合的(Sainis和Melzer 2005).PSI的E亚基是FNR在类囊体膜上的主要结合位点(Andersen等1992).这表明FNR的功能并不是简单的作为Calvin循环多酶复合体到PSI之间的膜连接器,FNR与GAPDH的结合可以认为是NADP/NADPH的通道,这样光反应产生的还原力NADPH就可更加便利的运输到暗反应,提高光合速率.总之,根据前人的研究可以推断,多酶复合体与类囊体膜结合可以使膜上的代谢物通道更加通畅,各类代谢途径可以互相连接,同时还可以用以解释高等植物中有效的光合电子传递和CO:固定之间的耦联.此外,非液相介质可以显着增加可溶性酶的稳定性和周转数量,酶与膜结合后会限制水在酶层中的分布,并在催化位点上产生一个非极性的环境,从而促使酶的稳定性和周转数量增加,进而更加便利的发挥其催化作用.酶与膜结合还可以延长酶的寿命,二者结合后可以阻止酶聚集和钝化(Bell等1995),因此,Calvin循环酶与类囊体膜结合的研究是有意义的.4结语Calvin循环在植物合成代谢中是一个独特的过程,在CO,同化中起作用,它与绿色植物,藻类和光合细菌的生物量和产量,温室气体减少等有着密切的联系.迄今对Calvin循环个体酶的研究比较多,长期以来一直认为Rubisco是这一途径的限速酶(Man1999),因此,对Rubisco在蛋白和基因水平上做了大量研究(Spreitzer和Salvucci2002).但Quick等(1992)对Rubisco是限速酶这一概念提出了质疑,Raines(2003)对Calvin循环的综述中也表明在各种环境条件下,Rubisco对碳固定的影响很小,FBPase,RPK,GAPDH,醛缩酶和转酮酶却有着不可忽视的作用,而SBPase是Calvin循环的限速酶,并且要更加重视对酶复合体的研究,特别是关于复合体中酶活性调控的机制的研究.因此这就要求我们重新考虑代谢途径的控制是否还将仍然停留在代谢途径单个酶的活性上..尽管Calvin—Benson循环多酶复合体已经得到证实,但仍有很多问题没有解决.首先是各个实验室得到的多酶复合体的组成有差异,不仅表现在不同物种上,同一物种之间也有很大的差异,这些差异需要用更多的材料作实验,这样才可在更大范围的高等植物叶绿体中进行比较,验证其普遍性;其次是揭示多酶复合体的空间结构特植物生理学通讯第4卷第3期,2008年6月397点,即各个酶是如何组装的,哪个酶在复合体的内部,哪个酶在复合体的外部,它们是如何相互作用的,其超分子的空间结构又是如何,迄今这方面的报道还很少,只有一些结构模型的猜想(图2),因此这也是未来研究Calvin循环多酶复合体可能要解决的问题;最后也是最本质的问题是多酶复合体的生理意义和其结构功能的研究,其目的都是为了揭示多酶复合体在生物体内的作用.因此在光合作用机制和蛋白质结构的研究中,多酶复合体的相互结合,并能表现出各个酶的催化活性的探讨,无论是在体内或是体外进行验证都有意义.参考文献AndersenB,SchellerHV,Moiler—LindbergB(1992).ThePSI—E subunitofPhotosYstemIbindsferredoxin:NADP oxidoreductase.FEBSLett.3ll:l69~l73?AndersonLE,GibbonsJT,WangXW(1996).Distributionoften enzymesofcarbonmetabolisminpea(Pisurnsativurn) chloroplasts.IntJPlantSci.157:525~538AvilanL,GonteroB,LebretonS,RicardJ(1997).Information transferinmultienzymecomplexes.2.TheroleofArg64of Chlarnydornonasreinhardtiiphosphoribulokinaseinthein—formationtransferbetweenglyceraldehydes一3一phosphatede—hydrogenaseandphosphoribu1okinase.EurJBiochem,250: 296~302BabadzhanovaMP,BabadzhanovaMA,AlievKA(2002).Freeand membrane—boundmultienzymecomplexeswithcalvincycle activitiesincottonleaves.RUSSJPlantPhysiol.49:592~597 BabadzhanovaMP,BabadzhanovaMA,AlievKA(2006).Devel—opmentalchangesintheactivitiesofenzymesoffreemul—tienzymecomplexesinvolvedintheBenson.Calvincycle. RUSSJPlantPhysiol.53:32~38BellG,HailingPJ,MooreBD,PatridgeJ,ReesDG(1995). Biocatalystsbebaviourinlowwatersystems.TrendsBiotech,l3:468~473BrendaSJW(2004).Metabolicchannelinginplants.AnnuRev PlantBiol,55:85~107GonteroB,CardenasML,RicardJ(1988).Afunctionalfive—enzymecomplexofchloroplastsinvolvedintheCalvincycle. EurJBiochem.173:437~443GonteroB,Giudici—OrtieoniM—T,RicardJ(1994).Themodula—tionofenzymereactionrateswithinmulti—enzyme rmationtransferwithinachloroplast multi—enzymecomplexcontainingribulosebisphosphate carboxylase—oxygenase.EurJBiochem,226:999~1006 GonteroB,MulliertG,RaultM,Giudici—OrticoniM—T.RicardJ (1993).Structuralandfunctionalpropertiesofamultien—zymecomplexfromspinachchloroplasts.2.Modulationof thekineticprope~iesofenzymesintheaggregatedstate.Eur JBiocbem.2l7:l075~l082GracietE,LebretonS,GonteroB(2004).Emergenceofnew regulatorymechanismsintheBenson—Calvinpathwayvia protein—proteininteractions:aglyceraldehyde一3一phosphate dehydrogenase/cPI2/J)hosphoribuIokinasecomplex.JExp Bot,55:1245~1254GracietE,GansP,WedelN,LebretonS,CamadroJM,GonteroB (2003).ThesmallproteinCP12:aproteinlinkerforsu—pramolecularcomplexassembly.Biochemistry,42:8l63~8170GracietE,LebretonS,CamadroJM,GonteroBf2002).Thermo—dynamicanalysisoftheemergenceofnewregulatoryprop—ertiesinaphOsphOribu1Okinase—glycera1dehyde3-phosphate dehydrogenasecomplex.JBiolChem,277:12697~12702 HermosoR,FelipeMR,VivoA.ChuecaA,LazaroJ,GorgeJL (1989).Immunogoldlocalizationofphotosyntheticfruc—tose一1,6-bisphOsphataseinpealeaftissue.PlantPhysiol,89:38l~385HermosoR,FonollaJ,deFelipeMR,VivoA,ChuecaA,LazaroJJ, Lopez—GorgeJ(1992).Doubleimmunogoldlocalizationof thioredoxinfandphotosyntheticfructose一1,6-bisphosphate inspinachleaves.PlantPhysiolBiochem,30:39~46HosurMV,SainisJK,KannanKK(1993).CrystallizationandX—rayanalysisofamultienzymecomplexcontainingRubisco andRuBP.JMolBio1.234:l274~l278HrazdinaG,JensenRA(1992).Spatialorganizationofenzymes inplantmetabolicpathways.AnnuRevPlantPhysiolPlantMolBio1.43:24l~267JebanathirajahJA,ColemanJR(1998).Associationofcarbonic anhydrasewithaCalvincycleenzymecomplexinNicotiana tabacurn.Planta.204:l77~l82KeletiT,OvadiJ,BatkeJ(1989).Kineticandphysico—chemical analysisofenzymecomplexesandtheirpossiblerolein controlofmetabolism.ProgBiophysMolBiol,53:105~152 LebretonS,GonteroB(1999).Memoryandimprintinginmultien—zymecomplexes.Evidenceforinformationtransfer fromglyceraldehydes一3-phosphatedehydrOgenasetO phosphoribulokinaseunderreducedstateinChlarnydorno—nasreinhardtii.JBiolChem,274:20879~20884LebretonS,GonteroB,AvilanL,RicardJ(1997).Information transferinmultienzymecomplex.1.Thermodynamicsof conformationalconstraintsandmemoryeffectsinthe bienzymeglyceraldehydes一3一phosphate—dehydrOgenase—phOsphOribulOkinasecomplexofChlarnydornonasreinhardtii chloroplasts.EurJBiochem,250:286~295MannCC(1999).Futurefood:bioengineer:geneticengineersaim tosoupupcropphotosynthesis.Science,283:314~316 MangeneyE,HawthornthwaiteAM,CoddC,GibbsSP(1987). ImmunocytochemicallocalizationofphOsphOribulOkinase incyanellesofCyanophorapardoxaandGlaucocystis nostochinearum.PlantPhysiol,84:1028~1032MarriL,TrostP,PupilloP,SparlaF(2005).Reconstitutionand propertiesoftherecombinantglyceraldehyde一3一phosphate dehydrogenase/CP12/phosphoribulokinasesupramolecular complexofArabidopsis.PlantPhysiol,139:1433~1443 MendiolaL,AkazawaT(1964).Partialpurificationandenzy—398植物生理学通讯第44卷第3期,2008年6月maticnatureoffractionIproteinofriceleaves.Biochemistry,3:174~179MoriH,TakabeT,AkazawaT(1984).Looseassociationof ribulose-l.5-biphosphatecarboxylasewiththylakoidmembranes.PhotosynthRes,43:17~28MoucheF,GonteroB,CallebautI,MornonJP,BoissetN(2002). Strikingconformationalchangesuspectedwithinthe phosphoribul0kinasedimmerinducedbyinteractionwith GAPDH.JBiolChem.277:6743,-6749MUllerB(1972).AlabileCO2-fixingenzymecomplexinspinach chloroplasts.ZNaturforsch.27B:925-932NicholsonS,EasterbyJS,PowlsR(1987).Propertiesofmultimeric proteincomplexfromchloroplastspossessingpotentialac—tivitiesofNADPHdependentg1yceraldehydes?3?phosphate dehydrogenaseandphosphoribulokinase.EurJBiochem.162: 423~431OvadiJ(1988).Oldpathway—newconcept:controlofglycolysis bymetabolite—modulateddynamicenzymeassociation. TrendsBiochemSci.13:486~490PerssonL—O,JohanssonG(1989).Studiesofprotein?protein interactionusingcountercurrentdistributioninaqueoustwo? phasesystems.PartitionbehaviorofsixCalvin?cycleen? zymesfromacrudespinach(Spinaciaoleracea)chloroplast extract.BiochemJ,259:863~870QuickWP,FichmerK.SchulzeE—D,WendlerR,LeegoogRC, MooneyH,RodermelSR,BogoradL,StittM(1992).De? creasedribulose—1.5?bisphosphatecarboxylase?oxygenasein transgenictobaccotransformedwithantisenserbcS.IV. Impactonphotosynthesisinconditionsofalterednitrogen supply.Planta,188:522~531RainesCA(2003).TheCalvincyclerevisited.PhotosynthRes,75: 1~10RaultM,Giudici—OrticoniM.T.GonteroB,RicardJ(1993). Structuralandfunctionalpropertiesofamulti??enzymecorn?? plexfromspinachchloroplasts.1.Stoichiometryofthe polypeptidechains.EurJBiochem.217:1065~1073RicardJ,Giudici?OrticoniM?T,GonteroB(1994).Themodula. tionofenzymereactionrateswithinmulti?enzyme complexes.1.Statisticalthermodynamicsofinformation transferthroughmulti?enzymecomplexes.EurJBiochem, 226:993~998SainisJK,HarrisGC(1986).Theassociationofribulose.1,5一bisphosphatecarboxylasewithphosphoriboisomeraseand phosphoribulokinase.BiochemBiophysResCommun,139: 947~954SainisJK.Harri?。
果糖16二磷酸酶

果糖-1,6-二磷酸酶(Fructose-1,6-bisphosphatase,简称FBPase)是一种催化反应的酶,它能够催化果糖-1,6-二磷酸向果糖-6-磷酸的转化,这是糖异生途径中的一个重要反应。
在糖异生途径中,FBPase参与到糖原的合成过程中。它的作用是将糖异生途径中的中间产物果糖-1,6-二磷酸转化为果糖-6-磷酸,这是糖异生途径中的最后一个步骤。这个反应需消耗ATP,同时也是一个可逆反应,可以被果糖-6-磷酸逆转。
可治性罕见病—果糖1,6一二磷酸酶缺乏症

可治性罕见病—果糖1,6一二磷酸酶缺乏症一、疾病概述果糖一1,6一二磷酸酶缺乏症(fructose -l,6- bisphosphatase deficiency)是一种罕见的遗传代谢病,呈常染色体隐性遗传。
目前尚不清楚其发病率,至今报道约100余例。
该病的致病基因是位于染色体9q22的FBP1基因。
当该基因发生突变,导致肝脏果糖一1,6一二磷酸酶缺乏或活性低下时,可造成1,6一二磷酸果糖转化为6-磷酸果糖的途径出现障碍,影响糖异生过程,造成果糖、乳酸、甘油等上游糖异生底物的堆积[1-3]。
二、临床特征该病的临床表现主要为发作性低血糖、高乳酸血症、代谢性酸中毒和酮症等,可致新生儿或婴儿死亡[4]。
发热、感染、进食不足及应激状态等常为本病发作的诱因。
本病也可在摄人大量果糖或蔗糖(1g/kg体重)后发作,静脉输注果糖或甘油有致命危险。
患儿发病年龄一般为生后1天~4岁。
由于乳糖由乳糖酶分解成葡萄糖和半乳糖的过程不受果糖一1,6一二磷酸酶的影响,婴儿常在断奶后才出现症状,发作期代谢紊乱较易纠正,而发作间期正常[5]。
最常见的首发症状为呕吐,其次为嗜睡、呼吸和心率增快等,血生化检查除低血糖、乳酸酸申毒外,还可伴有肝损害、凝血酶原时间延长等[6]。
三、诊断血生化检查显示低血糖、高乳酸血症、代谢性酸中毒,多伴有酮症。
发作期血、尿代谢筛查可发现糖异生底物堆积,可显示乳酸、3-羟基丁酸、甘油和3一磷酸甘油等的异常升高。
肝活检检测果糖一1,6一二磷酸酶的酶活性可确诊。
因肌肉组织中含有与肝脏不同的另外一种同工酶,本病患者在肌肉的活性并不下降,因此肌肉活检组织的酶活检测结果对本病无诊断价值。
在疾病的非发作期,甘油、果糖或丙氨酸负荷试验及禁食试验可诱发代谢紊乱,有助于疾病诊断,但并非确诊手段且存在一定危险,故不作为首选诊断方法。
仅在当临床及生化检测均高度支持本病,而基因检测和酶活性测定均正常的情况下,这种功能试验可尝试使用[5]。
磷酸果糖激酶研究进展

磷酸果糖激酶研究进展杨益坤生命科学学院SA16008074摘要:磷酸果糖激酶掌握着糖酵解开始的钥匙,是糖酵解中最重要的控制点。
糖酵解速度主要取决于磷酸果糖激酶,它是个限速酶,其受多个途径调控:受ADP别构激活、ATP别构抑制;受柠檬酸、脂肪酸别构抑制;受果糖-2,6-二磷酸的调控;受H+抑制。
生物体对磷酸果糖激酶的调控十分精细。
对于磷酸果糖激酶的研究和应用主要在于对于糖尿病的治疗,磷酸果糖激酶的变构效应的研究,磷酸果糖激酶在糖酵解中的地位等等。
本文就磷酸果糖激酶的研究进展进行综述。
关键词:磷酸果糖激酶别构激活别构抑制柠檬酸结合位点乳酸糖尿病地位一.磷酸果糖激酶简介名称:磷酸果糖激酶(fructose-1,6-bisphosphate)磷酸果糖激酶1(Phosphofructokinase-1;PFK-1)是一种糖酵解作用里一种重要的酶,是一种由4个次单位组成的异位(allosteric)酵素,可受多种活化剂与抑制剂调控。
糖酵解(glycolysis)是指在细胞质中将葡萄糖分解成为丙酮酸的过程,期间每分解一分子葡萄糖产生两分子丙酮酸以及两分子ATP。
属于呼吸作用的第一个阶段,随后根据有氧与否将丙酮酸进行柠檬酸循环或发酵。
在糖解作用里,PFK-1是负责将果糖-6-磷酸与ATP 转变成为果糖-1,6-双磷酸与ADP。
糖酵解过程中,6-磷酸果糖生成1,6-二磷酸果糖( Fructosel,6bisphosphate,F-1,6-BP),催化此反应的酶是6-磷酸果糖激酶1(6-phosphofructokinase1,PFK1),这是糖酵解途径的第二次磷酸化反应,需要ATP与Mg2+参与,ΔG"0=-14.2KJ/mol,反应不可逆。
6-磷酸果糖激酶1是糖酵解过程的主要限速酶,是糖酵解过程中的主要调节点。
至此,糖酵解完成了代谢的第一个阶段。
催化反应:果糖-6-磷酸+ATP→果糖-1,6-二磷酸+ADP辅助因子:Mg2+催化机制:镁离子将ATP外侧的两个磷酸基团作用,使磷原子更容易接受孤对电子的亲和进攻,使两个磷原子之间的氧桥所共用的电子对向氧原子一方转移,ATP的磷酸集团有利于氧桥断裂,并与果糖-6-磷酸分子结合,生成果糖-1,6-二磷酸。
果糖-1,6-二磷酸酶

果糖-1,6-二磷酸酶果糖-1,6-二磷酸酶(Fructose-1,6-bisphosphatase,简称为 FBPase)是一种参与葡萄糖代谢的酶,存在于几乎所有真核生物、许多原核生物以及一些病毒中。
它是Gluconeogenesis(糖异生)途径中的一个重要酶,负责将2分子果糖-1,6-二磷酸(FBP)水解为2分子磷酸果糖(F6P),从而使糖异生途径前进。
该酶是糖异生途径和糖原合成途径中的限速酶之一,在空腹时,肝脏中糖异生途径占主导地位,FBPase的活性是肝脏维持正常生理状态所必需的。
结构与功能FBPase 主要由 3 个结构域构成:N 端结构域、中间结构域和 C 端结构域,这些结构域在进化过程中具有高度的保守性。
N 端结构域含有磷酸化酶催化所需的脯氨酸残基,是酶催化和调节的关键区域。
中间结构域含有两个反向同构的催化中心,这些催化中心使 FBPase 能够高效地水解两个 FBP 分子,从而生成两个 F6P。
C 端结构域则对物质基础进行分类和整合,为 FBPase 酶学和生物学特性的整合提供基础。
FBPase 对许多环境因素,如 pH 值,离子强度和温度等,均表现出高度的敏感性。
稳定剂和抑制剂都能够显著地影响酶的活性。
对于细胞与机体内的 FBPase 来说,该酶的调节显得尤为重要。
急性调节通常由磷酸化和解磷酸化等化学修饰实现,而长期调节则主要由基因表达调控实现。
FBPase 的磷酸化和解磷酸化磷酸化和解磷酸化修饰对于 FBPase 的调节非常重要。
当细胞需要进入糖异生途径时,活性化的酶被磷酸化,并透过蛋白水解酶4的参与,磷酸化 FBPase 的活性在糖异生途径的起始阶段被激发,以产生更多的葡萄糖。
与此相反,当细胞需要停止进行糖异生途径时,F6P 是其调节的关键物质,彼时解磷酸化的 FBPase 更方便形成稳定的 F6P/FBP 催化活性平衡,从而使糖异生途径的前进效率下降。
改变 FBPase 的活性和构象调整代表了一种非常快速和可靠的方式,调整葡萄糖代谢产物的浓度,从而使细胞适应外界环境的需求。
1.6—二磷酸果糖临床研究介绍

1.6—二磷酸果糖临床研究介绍
潘世芬;安向光
【期刊名称】《医学理论与实践》
【年(卷),期】1994(007)001
【摘要】1.6—二磷酸果糖(fructose—1,6—Diphosphate)简称FDP。
它是一种具有调节机体糖代谢活性酶的作用,恢复及改善细胞分子水平代谢的药物。
近年研究表明,本药是治疗心力衰竭、急性心肌梗塞、心绞痛、休克、心肌炎等疾病的良好的辅助药物,且相对无副作用。
但临床应用上还有一定的局限性。
本文就有关国内外文献综述如下。
一、药理作用 FDP作用于细胞膜,激活磷酸果糖激酶(PEK),加速糖酵解。
【总页数】2页(P42-43)
【作者】潘世芬;安向光
【作者单位】不详;不详
【正文语种】中文
【中图分类】R972
【相关文献】
1.1.6二磷酸果糖联合黄芪注射液治疗病毒性心肌炎临床观察 [J], 刘文澜
2.参附注射液联合1.6-二磷酸果糖治疗小儿病毒性心肌炎的疗效观察 [J], 孙广斌
3.热敷缓解静脉输注1.6-二磷酸果糖(FDP)致痛的效果观察 [J], 段瑛;王建美
4.1.6—二磷酸果糖停跳液对心肌保护作用的临床研究 [J], 张国报;叶世铎
5.1.6二磷酸果糖联合黄芪注射液治疗病毒性心肌炎临床观察 [J], 刘文澜
因版权原因,仅展示原文概要,查看原文内容请购买。
果糖-1,6-二磷酸(FDP)含量检测试剂盒说明书__可见分光光度法UPLC-MS-4197
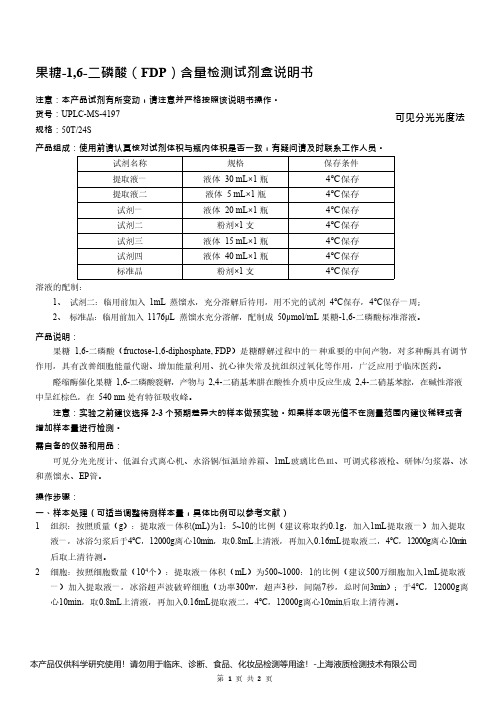
果糖-1,6-二磷酸(FDP)含量检测试剂盒说明书注意:本产品试剂有所变动,请注意并严格按照该说明书操作。
货号:UPLC-MS-4197可见分光光度法规格:50T/24S产品组成:使用前请认真核对试剂体积与瓶内体积是否一致,有疑问请及时联系工作人员。
试剂名称规格保存条件提取液一液体30mL×1瓶4℃保存提取液二液体5mL×1瓶4℃保存试剂一液体20mL×1瓶4℃保存试剂二粉剂×1支4℃保存试剂三液体15mL×1瓶4℃保存试剂四液体40mL×1瓶4℃保存标准品粉剂×1支4℃保存溶液的配制:1、试剂二:临用前加入1mL蒸馏水,充分溶解后待用,用不完的试剂4℃保存,4℃保存一周;2、标准品:临用前加入1176μL蒸馏水充分溶解,配制成50μmol/mL果糖-1,6-二磷酸标准溶液。
产品说明:果糖1,6-二磷酸(fructose-1,6-diphosphate,FDP)是糖酵解过程中的一种重要的中间产物,对多种酶具有调节作用,具有改善细胞能量代谢、增加能量利用、抗心律失常及抗组织过氧化等作用,广泛应用于临床医药。
醛缩酶催化果糖1,6-二磷酸裂解,产物与2,4-二硝基苯肼在酸性介质中反应生成2,4-二硝基苯腙,在碱性溶液中呈红棕色,在540nm处有特征吸收峰。
注意:实验之前建议选择2-3个预期差异大的样本做预实验。
如果样本吸光值不在测量范围内建议稀释或者增加样本量进行检测。
需自备的仪器和用品:可见分光光度计、低温台式离心机、水浴锅/恒温培养箱、1mL玻璃比色皿、可调式移液枪、研钵/匀浆器、冰和蒸馏水、EP管。
操作步骤:一、样本处理(可适当调整待测样本量,具体比例可以参考文献)1.组织:按照质量(g):提取液一体积(mL)为1:5~10的比例(建议称取约0.1g,加入1mL提取液一)加入提取液一,冰浴匀浆后于4℃,12000g离心10min,取0.8mL上清液,再加入0.16mL提取液二,4℃,12000g离心10mi n 后取上清待测。
果糖1,6-二磷酸盐

果糖1,6-二磷酸盐
果糖1,6-二磷酸盐(fructose-1,6-bisphosphate)是一种生物分子,是糖酵解途径中的重要中间产物。
它的分子式为C6H14O12P2,分子量为340.116 g/mol。
果糖1,6-二磷酸盐在糖酵解途径中的作用是将葡萄糖分解成两个三碳分子的丙酮酸。
它是糖酵解途径中的关键中间产物,也是糖酵解途径中ATP生成的重要来源。
在糖酵解途径的第三步,果糖1,6-二磷酸盐被酶类催化分解成为两个三碳分子的丙酮酸和磷酸二酯。
果糖1,6-二磷酸盐的研究历史可以追溯到20世纪初。
1905年,德国生物化学家Emil Fischer首次发现了果糖1,6-二磷酸盐的存在,并研究了它在糖酵解途径中的作用。
此后,随着生物化学和分子生物学的发展,人们对果糖1,6-二磷酸盐的研究也越来越深入。
近年来,果糖1,6-二磷酸盐在医学和生物技术领域的应用也越来越广泛。
例如,研究人员利用果糖1,6-二磷酸盐的特殊性质,开发出了一种新型的生物传感器,可以用于检测血液中的葡萄糖浓度。
此外,果糖1,6-二磷酸盐还被广泛应用于生物制药和生物能源等领域。
总之,果糖1,6-二磷酸盐是糖酵解途径中的重要中间产物,具有重要的生物学意义和应用前景。
果糖 1,6- 二磷酸醛缩酶(Fructose 1,6 bisphosphate aldolase ,FDA )试剂盒说明书

货号:MS2210 规格:100管/96样果糖 1,6-二磷酸醛缩酶(Fructose 1,6 bisphosphate aldolase,FDA)试剂盒说明书微量法注意:正式测定之前选择 2-3个预期差异大的样本做预测定。
测定意义:植物叶绿体中果糖1,6-二磷酸醛缩酶是光合作用中参与calvin循环的重要酶。
催化果糖1,6-二磷酸和景天庚酮糖1,7-二磷酸的合成反应,在各种逆境胁迫下表现不同的响应。
测定原理:果糖1,6-二磷酸醛缩酶催化果糖1,6-二磷酸生成3-磷酸甘油醛和磷酸二羟丙酮,在磷酸丙糖异构酶和α-磷酸甘油脱氢酶作用下催化NADH和磷酸二羟丙酮生成NAD和α-磷酸甘油,340nm处吸光值的变化可反映果糖1,6-二磷酸醛缩酶活性的高低。
自备实验用品及仪器:天平、震荡仪、低温离心机、研钵、紫外分光光度计/酶标仪、微量石英比色皿/96孔板。
试剂组成和配制:提取液一:液体100mL×1瓶,4℃保存。
提取液二:液体100mL×1瓶,4℃保存。
试剂一:液体10mL×1瓶,4℃避光保存。
试剂二:粉剂×1瓶,-20℃避光保存。
临用前加2mL蒸馏水充分溶解;用不完的试剂分装后-20℃保存,禁止反复冻融。
试剂三:粉剂×1瓶,-20℃避光保存。
临用前加2 mL蒸馏水充分溶解;用不完的试剂分装后-20℃保存,禁止反复冻融。
试剂四:粉剂×1瓶,-20℃避光保存。
临用前加2 mL蒸馏水充分溶解;用不完的试剂分装后-20℃保存,禁止反复冻融。
试剂五:液体2mL×1瓶,4℃避光保存。
酶液提取:①总FDA酶提取:建议称取约0.1g样本,加入1mL提取液一,冰浴匀浆后超声破碎(冰浴,200W,破碎3s,间歇7s,总时间1min),然后4℃,8000g离心10min,取上清测定。
②胞浆和叶绿体FDA酶的分离:按照植物组织质量(g):提取液体积(mL)为1:5~10的比例(建议称取约0.1g样本,加入1mL提取液一),冰浴匀浆后于4℃,200g离心5min,弃沉淀,取上清在4℃,8000g离心10min,取上清用于测定胞浆FDA酶活性,取沉淀加1mL提取液二,震荡溶解后超声破碎(冰浴,200W,破碎3s,间歇7s,总时间1min),然后4℃,8000g离心10min,取上清测定叶绿体中FDA酶活性。
丙糖磷酸异构酶_果糖_1_6_二磷酸醛缩酶及果糖_1_6_二磷酸酶的共表达

ISSN 058229879生物化学与生物物理学报ACTA BIOCHIMICA et BIOPHYSICA SINICA 2001,33(1):131-136CN 3121300/Q收稿日期:2000206229 接受日期:2000209208中国科学院基础性研究重点资助项目3联系人:Tel ,02126416330021429;Fax ,021*********;e 2mail ,hbchen @研究简报丙糖磷酸异构酶、果糖21,62二磷酸醛缩酶及果糖21,62二磷酸酶的共表达唐功利 杨春松 鲍建绍 王燕芳 陈海宝3(中国科学院上海有机化学研究所生命有机化学国家重点实验室,上海200032)施定基 刘凤龙(中国科学院植物研究所光合室,北京100093)摘要 致力于建立一条控制或降低大气中CO 2浓度的途径,选择对蓝藻进行代谢工程以便改进其光合固定CO 2的效率。
作为研究的初始阶段,将编码丙糖磷酸异构酶、果糖21,62二磷酸醛缩酶及果糖21,62二磷酸酶的3个基因构建进一个由启动子t rc 控制的表达质粒p TrcFA T ,成功地在大肠杆菌中实现了上述3个基因的活性共表达。
活性测定结果显示:从1L 培养液获得的破菌上清液每分钟可以催化二羟丙酮磷酸(DHAP )转化成700μmol 果糖262磷酸。
在此基础上进一步构建了这3个基因共表达的大肠杆菌2蓝藻穿梭表达质粒,也在大肠杆菌中实现了活性表达。
当外源基因的操纵子与载体质粒以大于1∶1的比例进行构建时,可显著提高外源基因的表达量及相应的酶活性。
关键词 丙糖磷酸异构酶;果糖21,62二磷酸醛缩酶;果糖21,62二磷酸酶;共表达 大气中温室气体浓度上升导致的全球气温变暖、气候异常已引起人们的普遍关注,其中CO 2是主要的温室气体[1]。
按照政府间气候变化专业委员会(IPCC )的分析,若按照目前CO 2排放的趋势增长,到下个世纪,全球平均温度将以每10年0.2~0.4℃的速度升高[2],从而带来气候变暖、海平面升高、耕地减少、农业病虫害加剧、北极圈CO 2生态失衡等一系列恶果。
磷酸戊糖途径-

(5) 二分子 五碳糖得基 团转移反应
H C OH
CH2OPO3H2
核糖-5-磷酸
ribose 5-phosphate
转酮反应
CH2OPO3H2
景天庚酮糖-7-磷酸
sedoheptulose 7-phosphate
(6)七碳糖与三碳糖得基团转移反应
CH2OH CO
Mg2+或 Mn2+
CHO H C OH
CH2OH CO HO C H
H C OH
H C OH
CH2OPO3H2
果糖-6-磷酸
fructose 6-phosphate
(7)四碳糖与五碳糖得基团转移反应
CH2OH
CHO
转酮反应
CO
H C OH
CH2OH
HO C H H C OH
CHO
CH2OPO3H2
甘油醛-3-磷酸
CO
glyceraldehyde 3-phosphate HO C H
总反应图 反应式 转酮醇酶与转醛缩酶比较 特点
大家有疑问的, 可以询问和交流
可以互相讨论下, 但要小声点
6NADPH 6CO2 6×Ru5P
磷酸戊糖途径:
2×Xu5P 2×R5P
6×6-磷酸 葡萄糖酸
2×S7P
2×GAP
6H2O
6×6-磷酸葡 萄糖酸内酯
2× Xu5P
6NADPH
2×GAP
6×葡萄糖-6-磷酸
核酮糖-5-磷酸
ribulose 5-phosphate
6-磷酸葡萄糖酸脱氢酶
6-phosphogluconate dehydrogenase
(4) 三种五碳糖得互换:
1,6-二磷酸果糖的脑保护作用研究进展(精)

1,6-二磷酸果糖的脑保护作用研究进展袁宝莉,宋晓红,李瑞林(西安交通大学第二医院儿科,陕西西安710004)摘要1,6-二磷酸果糖是葡萄糖分解代谢的产物,近年来被广泛用于心、肾、肝、脑的缺血性治疗。
该文综述了1,6-二磷酸果糖的脑保护作用机制剂量范围及其应用,探讨对临床缺血性脑疾患的治疗意义。
关键词1,6-二磷酸果糖;钙离子;缺血性脑损伤1,6-二磷酸果糖(fructose-1 , 6- dip hosphate, FDP;or fructose-1 ,6-bisphosphate , FBP)是葡萄糖分解代谢的中间产物,在体外经静脉注射后可广泛分布于心、肾、肝、脑等器官。
自上世纪70年代开始,FDP就被广泛应用于心脏骤停、心肌梗死、心肌缺血及肾脏缺血等的治疗上[ 1,2 ]。
但直到80年代后期,研究人员才发现FDP可以通过血脑屏障被中枢神经系统代谢,并且它对于脑功能的保护和恢复具有独特作用,于是学术界开始了对FDP应用于脑血管疾病的实验性研究。
这为目前尚未有特效治疗的缺血性脑损伤开拓了一条新思路。
1. 1,6-二磷酸果糖的作用机制学术界对于FDP的脑保护作用机制一直存在三种假设。
第一种由Markov等(1981年)提出,认为机体将摄入的FDP作为一种高能物质及糖酵解的调节剂,以此来加强无氧代谢;第二种由Galzigna等(1989年)提出,认为FDP并未进入到细胞内发挥作用,它通过改变离子流动包括氢离子(H+)流动来调节细胞内PH值;第三种说法(1992年)认为FDP通过鳌合血清中游离钙离子(Ca 2+ )来减少进入细胞内的Ca2+ 譬如FDP可以减少心肌组织Ca2+ 摄入来减少游离Ca2+ ;此外尚存在有其他一些假说,不一一详述。
近年来学者们在上述假说的基础上,运用体外细胞培养技术及其他高科技手段对FDP的作用机制进行探索,已取得一定进展。
1 .1稳定细胞内Ca2+ ,维持内环境稳定Ca2+ 内流是细胞破环的主要原因,FDP通过减少由缺氧导致的神经元胞浆内的Ca2+ 上调来发挥神经保护作用[ 3 ] 。
翘嘴红鲌(Erythroculter ilishaeformis)果糖-1,6-二磷酸醛缩酶(ALDO-C)基因定位

第50卷 第5期 海 洋 与 湖 沼Vol.50, No.5 2019年9月OCEANOLOGIA ET LIMNOLOGIA SINICASep., 2019* 浙江省农业(水产)新品种选育重大科技专项, 2016C02055-1号。
王志刚, 硕士研究生, E-mail: 623474593@ ① 通信作者: 郑荣泉, 博士, 教授, E-mail: zhengrq@ 收稿日期: 2019-03-10, 收修改稿日期: 2019-04-14翘嘴红鲌(Erythroculter ilishaeformis )果糖-1, 6-二磷酸醛缩酶(ALDO-C )基因定位、克隆及表达分析*王志刚1 刘士力4 李贤露1 吕卓云1 毛丽盈1 唐超然1 郑荣泉1, 2, 3①(1. 浙江师范大学化学与生命科学学院 金华 321004; 2. 浙江省野生动物生物技术与保护利用重点实验室 金华 321004; 3. 浙江师范大学行知学院 金华 321004; 4. 浙江省淡水水产研究所 农业农村部淡水渔业健康养殖重点实验室/浙江省淡水水产遗传育种重点实验室 湖州 313001)摘要 鲌翘嘴红(Erythroculter ilishaeformis )鲌是大型淡水鱼类鳊亚科(Abramidinae)中最大的一种鱼, 具有较高的经济价值。
但其饲料转化率及抗病性研究相对较少, 相关基因信息缺乏。
本研究以翘鲌嘴红为对象, 利用RACE 鲌技术克隆翘嘴红果糖-1, 6-二磷酸醛缩酶(ALDO-C )基因, 该基因cDNA 全长1945bp, 其中ORF 区975bp, 编码325个氨基酸, 5′非编码区933bp, 3′非编码区37bp 。
通过实时荧光定量PCR 检测了ALDO-C 在不同组织中相对表达水平。
发现ALDO-C 鲌基因在翘嘴红的肾、肝、肌肉、性腺中均有表达, 且在肾中表达量最高, 显著高于其他组织。
同时采用石蜡切片、H.E 染色和原位杂交染色, 鲌观察分析翘嘴红的肾组织显微结构和ALDO-C 基因的表达定位, 结果表明, ALDO-C 在鱼类肾单位和集合管结构、调节肾组织水平衡及抗病性方面有重要作用。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
植物叶绿体中果糖1,6-二磷酸醛缩酶试剂盒使用说明
分光光度法注意:正式测定之前选择2-3个预期差异大的样本做预测定。
货号:BC2000
规格:50管/48样
产品内容:
提取液一:液体50mL×1瓶,4℃保存。
提取液二:液体25mL×1瓶,4℃保存。
试剂一:液体25mL×1瓶,4℃避光保存。
试剂二:粉剂×1瓶,-20℃避光保存。
临用前加5mL蒸馏水充分溶解。
试剂三:粉剂×1瓶,-20℃避光保存。
临用前加5mL蒸馏水充分溶解。
试剂四:粉剂×1瓶,-20℃避光保存。
临用前加5mL蒸馏水充分溶解。
试剂五:液体×1瓶,4℃避光保存。
产品说明:
植物叶绿体中果糖1,6-二磷酸醛缩酶是光合作用中参与calvin循环的重要酶。
催化果糖1,6-二磷酸和景天庚酮糖1,7-二磷酸的合成反应,在各种逆境胁迫下表现不同的响应。
果糖1,6-二磷酸醛缩酶催化果糖1,6-二磷酸生成3-磷酸甘油醛和磷酸二羟丙酮,在磷酸丙糖异构酶和α-磷酸甘油脱氢酶作用下催化NADH和磷酸二羟丙酮生成NAD和α-磷酸甘油,340nm处吸光值的变化可反映果糖1,6-二磷酸醛缩酶活性的高低。
自备仪器和用品:
天平、低温离心机、震荡仪、研钵、紫外分光光度计、1mL石英比色皿。
操作步骤:
一、酶液提取
按照植物组织质量(g):提取液体积(mL)为1:5~10的比例(建议称取约0.1g,加入1mL提取液一)加入提取液一,冰浴匀浆后于4℃,1000g离心5min,取上清在4℃,4500g离心20min,去上清,取沉淀加0.5mL提取液二,置于震荡仪上震荡30s溶解,置冰上30min,重复操作一次(或者震荡溶解后超声,超声条件为功率200W,破碎3s,间歇7s,总时间1min),然后4℃,12000g离心10min,取上清待测。
二、测定操作
1.分光光度计预热30min,调节波长至340nm,蒸馏水调零。
2.取1mL石英比色皿,依次加入500μL试剂一,100μL试剂二,100μL试剂三,
100μL试剂四,100μL粗酶液,充分混匀,记录340nm处10s的吸光值A1和310s的吸光值A2,△A=A2-A1
三、计算公式:
(1)按照样本蛋白浓度计算
酶活单位定义:每毫克组织蛋白每分钟消耗1nmol的NADH定义为一个酶活力单位。
FDA(nmol/min/mg prot)=ΔA÷(ε×d)×V反总÷(V样×Cpr)÷T=321.54×ΔA ÷Cpr
(2)按照样本质量计算
酶活单位定义:每克组织每分钟消耗1nmol的NADH定义为一个酶活力单位。
FDA(nmol/min/g)=ΔA÷(ε×d)×V反总÷(W×V样÷V样总)÷T=321.54×ΔA ÷W
(3)按照细胞数量计算
酶活单位定义:每104个细胞每分钟消耗1nmol的NADH定义为一个酶活力单位。
FDA(nmol/min/104cell)=ΔA÷(ε×d)×V反总÷(V样×细胞数量÷V样总)÷T =321.54×ΔA÷细胞数量
(4)按照液体体积计算
酶活单位定义:每毫升液体每分钟消耗1nmol的NADH定义为一个酶活力单位。
FDA(nmol/min/mL)=ΔA÷(ε×d)×V反总÷V样÷T=321.54×ΔA
V反总:反应体系总体积,1mL;ε:NADH摩尔消光系数,6.22×103L/mol/cm;d:比色皿光径,1cm;V样:加入样本体积,0.1mL;V样总:加入提取液体积,1mL;T:反应时间,5min;Cpr:样本蛋白质浓度,mg/mL;W:样本质量,g
注意事项:
配置好的试剂二、试剂三、试剂四3天内使用完。