重要养殖鱼类功能基因组和分子设计育种的基础研究
分子生物学技术在水产动物中的研究与应用

分子生物学技术在水产动物中的研究与应用分子生物学技术在水产动物中的研究与应用一、简介水产动物是指以水为生活环境的动物,主要包括鱼类、虾类、贝类、藻类等,是一类重要的人类食物资源,也是海洋生态系统中的重要组成部分。
近年来,随着分子生物学技术的发展,在水产动物中分子生物学的研究也有所增加。
分子生物学技术可以帮助我们了解水产动物的基因、基因组、表观遗传学等,并且能够帮助我们更好地利用水产动物,如用于育种、克隆、营养素提取等。
本文将从水产动物基因组学研究、水产动物遗传育种、水产动物克隆、水产动物营养素提取四个方面介绍分子生物学在水产动物中的研究与应用现状。
二、分子生物学技术在水产动物基因组中的应用水产动物的基因组学研究是利用分子生物学技术研究不同水生生物的基因组结构和组成的一种研究方法。
近年来,在水产动物的基因组研究中,有许多研究都利用分子生物学技术来揭示基因组结构和组成。
例如,黑鲷鱼(C.melanurus)基因组已被全面测序,湖茈蝦(C.aquaticus)的基因组也已被测序,报道完整的基因组结构和全基因组的组成分析。
此外,蜗牛(B.scabricus)的基因组也已被测序,报道了基因组的完整结构和组成。
三、分子生物学技术在水产动物遗传育种中的应用利用分子生物学技术进行水产动物遗传育种,可以准确检测合适的水产动物种类和品种,为水产品质量的改良和提高提供有力的理论依据和技术支持。
目前,在水产动物遗传育种中应用的分子生物学技术主要有一代测序技术,核酸杂合子检测技术,以及近似群体分析技术等。
四、分子生物学技术在水产动物克隆中的应用分子克隆技术是一种利用外源质粒中的特定基因进行动物克隆的技术。
近年来,随着分子克隆技术的发展,许多水产动物都已被克隆,如章鱼(M.octopus)、黄玉米虾(P.monodon)、鳗鱼(M.shiloi)等。
利用分子克隆技术可以大大提高水产动物的产量,为水产养殖提供新的发展方向。
五、分子生物学技术在水产动物营养素提取中的应用随着人类饮食习惯的改变,对水产动物中的营养素的提取也越来越受到重视,如蛋白质、脂肪、矿物质、氨基酸、维生素等。
2010年973项目

2010CB227100 高效规模化太阳能热发电的基础研究 2010CB227200 大规模高效液流电池储能技术的基础研究 2010CB227300 多能源互补的分布式冷热电联供系统基础研究
1
序号
项目编号
Байду номын сангаас
项目名称 东南大学
承担单位
首席科学家 洪伟 刘新宇 任晓敏 许宁生 刘铁根 李波 孙家广 蒋昌俊 陈章渊 余少华 夏军 杨修群 吕达仁 周名江 石建省 赵美训 王超
超高频、大功率化合物半导体器件与集成技术基 础研究 新型光电子器件中的异质兼容集成与功能微结构 2010CB327600 体系基础研究 新型微显示和场发射平板显示高品质化及应用的 2010CB327700 基础研究
1315.00 1328.00 1312.00 1345.00 1331.00 1440.00 1373.00 1418.00 1409.00 1379.00 1167.00 1184.00 1290.00 1433.00 1268.00 1317.00 1195.00
深部重大工程灾害的孕育演化机制与动态调控理 论
2010CB732100 城市地下工程安全性的基础理论研究 2010CB732200 生物质转化为高值化材料的基础科学问题 2010CB732300
清洁能源生产和环境治理中稀土催化材料应用的 基础研究
2010CB732400 仿生分子识别技术在生物医学应用的基础研究
项目编号
项目名称 钢铁研究总院 中南大学 西安交通大学
承担单位
首席科学家 董瀚 邱冠周 孙军 益小苏 孙晓峰 朱敏 张文军 李世海 刘日平 李晓谦 戴琼海 吴一戎 冯夏庭 张顶立 孙润仓 卢冠忠 鞠熀先
2009年新立项973及重大研究计划项目
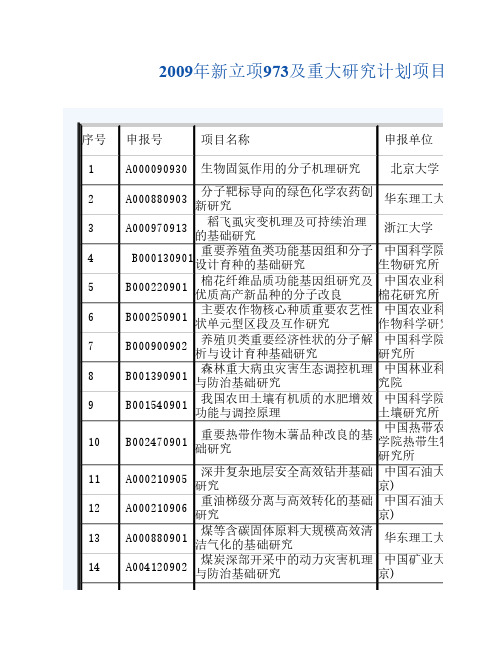
84 85 86 87 88 89 90 91 92 93 94 95 96 97 98
B000280901 应用 B000710904 B001110901 B001670901 B002950901 B007170901 A000080937 A000090941 A000300903 A000510901 A000980901 重大工程地质灾害的预测理论及 数值分析方法研究 城市环境要素高分辨率遥感谱像 合一认知理论与方法 稀疏微波成像的理论、体制和方 法研究 深部重大工程灾害的孕育演化机 制与动态调控理论 心脑血管易损斑块的高分辨成像 识别与风险评估预警体系重大问 题的基础研究 生物膜动态变化的分子机理与功 能研究 攻击与亲和社会行为的机理和异 常:多学科多层次交叉研究 基于精密测量物理的引力及相关 物理规律研究 复杂装备研发数字化工具中的计 算力学和多场耦合若干前沿问题 海相烃源岩形成的地球生物学过 程 抗体-抗原分子识别的结构基础 和功能研究
中国电力科学研 究院 中国科学院电工 研究所 中国科学院大连 化学物理研究所 中国科学院工程 热物理研究所 清华大学 北京大学 北京邮电大学 北京邮电大学 华中科技大学 东南大学 中山大学 天津大学 北京航空航天大 学 同济大学 中国科学院数学 与系统科学研究院 中国科学院微电 子研究所 武汉邮电科学研 究院 河海大学
银屑病的系统生物学研究 心脏间隔缺损形成、发展和干预 的基础研究
炎症过程中细胞间相互作用的信 号转导机制及其应用研究 恶性肿瘤发生及其早期防治的基 础研究 基于系统生物医学基础的白血病 临床转化研究 利用遗传地理时空模型预测H5N1 禽流感病毒的传播及控制策略研 清华大学 究 重要人兽共患胞内寄生菌病流行 石河子大学 特征及病原致病机制研究 中国科学院武汉 重要病毒的入侵机制研究 病毒研究所 我国重要食源性寄生虫病的发病 中山大学 机制及防治研究 基于“肾藏精”的脏象理论基础 上海中医药大学 研究 经脉体表特异性联系的生物学机 广州中医药大学 制及针刺手法量效关系的研究 以量-效关系为主的经典名方相 中国中医科学院 关基础研究 广安门医院 光电功能晶体结构性能、分子设 计、微结构设计与制备过程的研 南京大学 究 微生物冶金过程强化的基础研究 中南大学
973计划规划2004年度立项项
附件:973计划2004年度立项项目、项目首席科学家、项目依托部门策划书策划书活动策划书模板一、策划书名称尽可能具体的写出策划名称,如“×年×月×日信息系×活动策划书”,置于页面中央。
策划书二、活动背景:这部分内容应根据策划书的特点在以下项目中选取内容重点阐述具体项目有:基本情况简介、主要执行对象、近期状况、组织部门、活动开展原因、社会影响、以及相关目的动机。
三、活动目的及意义:活动的目的、意义应用简洁明了的语言将目的要点表述清楚;在陈述目的要点时,该活动的核心构成或策划的独到之处及由此产生的意义都应该明确写出。
四、活动名称:根据活动的具体内容影响及意义拟定能够全面概括活动的名称。
五、活动目标:此部分需明示要实现的目标及重点(目标选择需要满足重要性、可行性、时效性)。
六、活动开展作为策划的主题部分,表述方面要力求详尽,不仅仅局限于用文字表述,也可适当加入统计图表、数据等,便于统筹。
活动开展应包括活动流程安排、奖项设置、时间设定等。
涉及到奖项评定标准、活动规则的内容可选择以附录的形式出现。
活动流程安排大致可以分为三个阶段:(一)活动准备阶段(包括海报宣传、前期报名、赞助经费等);(二)活动举办阶段(包括人员的组织配置、场地安排情况等);策划书注:须注明开展活动的阶段负责人、指导单位、参加人数等信息。
(三)活动后续阶段(包括结果公示、活动开展情况总结等);注:如有涉及校园卫生、环境等情况,应及时清理。
七、经费预算:活动的各项费用在根据实际情况进行具体、周密的计算后,用清晰明了的形式列出。
八、活动中应注意的问题及细节:内外环境的变化,不可避免的会给方案的执行带来一些不确定性因素,因此,当环境变化时是否有应变措施,损失的概率是多少,造成的损失多大等也应在策划中加以说明。
九、活动负责人及主要参与者:注明组织者、参与者姓名、单位(如果是小组策划应注明小组名称、负责人)。
973项目申报书——重要养殖鱼类功能基因组和分子设计育种的基础研究精编版

项目名称:重要养殖鱼类功能基因组和分子设计育种的基础研究首席科学家:桂建芳中国科学院水生生物研究所起止年限:2010年1月-2014年8月依托部门:中国科学院湖北省科技厅一、研究内容基于拟解决的鱼类分子设计育种的策略和理论基础以及鱼类分子设计育种的可行性途径这两个关键科学问题,本项目拟集中在鱼类主要经济性状如生殖、生长、抗性的功能基因组和分子设计育种的基础方面,重点解析这些主要经济性状的基因调控网络,开拓关键技术,其主要研究内容包括:1)鱼类生殖基因调控网络及其分子设计育种的基础研究采用基因转移、morpholino介导的基因敲降等技术,配合整体原位杂交、细胞示踪和免疫荧光定位等方法,重点解析鱼类生殖质(germ plasm)的基因调控网络;揭示重要生殖调控基因在鱼类卵母细胞成熟和卵-胚转换中的功能作用及其调控机制;探讨鱼类配子发生过程中双亲特异性甲基化在生殖调控中的作用及其及其调控网络;通过大规模RFLP分析等呈现技术,筛选雌雄个体间性别特异的DNA片段标记,进而通过Genomic Walking克隆分离与性别决定和性别分化相关基因;揭示鱼类性别决定和性别分化的基因调控网络及其作用机理;筛选鉴定出可用于鱼类分子设计育种的性别特异表达基因和分子标记;开拓生殖调控基因和分子标记用于鱼类分子设计育种的可行性途径。
2)鱼类生长的基因调控网络和分子设计育种的基础研究采用morpholino介导的基因敲降、整体原位杂交、细胞示踪、免疫荧光定位以及组织碎片灌流和细胞孵育等在体和离体研究等技术,重点解析鱼类下丘脑/垂体控制生长的基因调控网络;揭示鱼类性成熟和生长的相互协调及其调控机理;探讨脑肠肽/生长激素/类胰岛素生长因子信号通路及其对鱼类生长的调控机制;揭示卵泡抑素等抑肌素相关基因在调控鱼类肌肉细胞增殖和鱼肉蛋白/脂肪平衡的作用机理;研制可控不育性转卵泡抑素等基因的转基因鱼品系;阐明鱼类个体大小调控基因的系统发育和进化规律;开拓生长基因调控网络和分子标记尤其是主控基因用于鱼类分子设计育种的可行性途径。
牙鲆sox9基因的克隆与表达研究

第29卷第1期2011年1月海洋科学进展ADVANCES IN MARINE SCIENCE Vol.29No.1Januar y,2011牙鲆sox9基因的克隆与表达研究*文爱韵1,2,尤锋1,*,孙鹏1,徐冬冬1,吴志昊1,马得友1,李军1,张培军1(1.中国科学院海洋研究所海洋生物学重点实验室,山东青岛266071; 2.中国科学院研究生院,北京100039)摘要:通过Genome walking 方法克隆得到了牙鲆(P a r alichthys olivaceus)sox9的基因组全长序列,对其在成鱼各组织的差异表达进行了RT -PCR 分析。
结果显示:牙鲆sox9基因全长3573bp,包含3个外显子和2个内含子,编码的蛋白全长478aa,含有高度保守的H MG 结构域;sox9基因在性腺的表达具有两性差异,精巢中的表达明显强于卵巢中的表达,属1个性别相关基因,但其在肝脏、心脏、鳃、脑、眼和胃等多种组织中也有不同程度的表达。
据此进一步分析和讨论等sox9基因在牙鲆性腺发育中的作用。
关键词:牙鲆(P a ra lichthys olivaceus);sox9;基因克隆;两性差异表达中图分类号:Q344文献标识码:A 文章编号:1671-6647(2011)01-0097-08sox9基因属于sox 基因家族,其编码的蛋白质在H MG(H igh mobility group box)保守盒内与哺乳类雄性性别决定基因sr y 的相似性大于60%[1],故该基因也应与性别相关。
研究表明,哺乳类sox9基因发生突变后有75%的雄性个体发育为中性或雌性。
该基因受到其上游的sr y 的转录调节[2],并在性腺中调控着下游基因的表达。
如sox9下调wnt4(wingless -type MMT V integration site family,member 4)的表达,性腺将发育成睾丸[3];sox9也可以参与激活amh(ant-i Mullerian hor mone)[4],使哺乳动物性腺向着精巢方向发育[5]。
科技部关于发布国家重点基础研究发展计划(含重大科学研究计划)2018年结题项目验收结果的通知

科技部关于发布国家重点基础研究发展计划(含重大科学研究计划)2018年结题项目验收结果的通知文章属性•【制定机关】科学技术部•【公布日期】2019.09.09•【文号】国科发基〔2019〕308号•【施行日期】2019.09.09•【效力等级】部门规范性文件•【时效性】现行有效•【主题分类】科技计划正文科技部关于发布国家重点基础研究发展计划(含重大科学研究计划)2018年结题项目验收结果的通知国科发基〔2019〕308号各有关项目依托部门:按照《国家重点基础研究发展计划管理办法》和《国家重点基础研究发展计划专项经费管理办法》有关规定,科技部组织完成了国家重点基础研究发展计划(973计划)2013年立项的3个项目、2014年立项的167个项目的结题验收。
现将项目验收结果通知如下。
1.“超强激光驱动粒子加速及其重要应用”等170个项目自立项实施以来,总体执行情况较好,达到了预期目标,予以通过验收。
其中,“大型飞机电液动力控制与作动系统新体系基础研究”等51个项目验收结果为优秀,“大型风力机的关键力学问题研究及设计实现”等119个项目验收结果为良好。
2.“农作物重要病毒病昆虫传播与致害的生物学基础”等170个项目财务验收结果为通过财务验收。
对于课题结余资金的处理,科技部将按照财政科研项目资金管理的有关规定执行。
特此通知。
附件:973计划(含重大科学研究计划)2018年结题项目验收结果科技部2019年9月9日附件973计划(含重大科学研究计划)2018年结题项目验收结果项目编号项目名称项目首席科学家项目第一承担单位项目依托部门项目验收结果财务验收结果973计划农业科学等9个领域2013CBA01500超强激光驱动粒子加速及其重要应用张杰上海交通大学教育部、上海市科学技术委员会优秀通过2013CBA01600与硅技术融合的石墨烯类材料及其器件的研究高鸿钧中国科学院物理研究所中国科学院优秀通过2013CBA01700波的衍射极限关键科学问题研究罗先刚中国科学院光电技术研究所中国科学院优秀通过2014CB046200大型风力机的关键力学问题研究及设计实王同光南京航空航天大学江苏省科学技术厅良好通过现2014CB046300深部危险煤层无人采掘装备关键基础研究葛世荣中国矿业大学江苏省科学技术厅、教育部良好通过2014CB046400大型飞机电液动力控制与作动系统新体系基础研究焦宗夏北京航空航天大学工业和信息化部优秀通过2014CB046500大型航空复合材料承力构件制造基础贾振元大连理工大学教育部优秀通过2014CB046600新一代超大型运载火箭薄壁结构制造的科学问题来新民上海交通大学教育部良好通过2014CB046700高服役性能海洋动力定位装备制造的基础研究邵新宇华中科技大学湖北省科学技术厅、教育部良好通过2014CB046800深海水下油气输送系统安全运行与风险控制余建星天津大学教育部、天津市科学技术委员会良好通过2014CB046900深部复合地层围岩与TBM的相互作用机理及安全控制刘泉声武汉大学湖北省科学技术厅、教育部良好通过2014CB047000山区支线机场高填方变形和稳定控制关键基础问题研究姚仰平北京航空航天大学工业和信息化部良好通过2014CB047100重大岩体工程灾害模拟、软件及预警方法基础研究唐春安大连理工大学教育部良好通过2014CB049000计及缺陷敏感性的网格加筋筒壳结构轻量化设计理论与方法王博大连理工大学辽宁省科学技术厅优秀通过2014CB049100高速铁路软土路基长期运营沉降与环境振动控制冯世进同济大学教育部、上海市科学技术委员会良好通过2014CB049400超深井大型提升装备设计制造及安全运行的基础研究邹声勇中信重工机械股份有限公司河南省科学技术厅良好通过2014CB049500大面积光栅纳米精度制造中的基础问题研究唐玉国长春奥普光电技术股份有限公司吉林省科学技术厅优秀通过2014CB138100小麦产量和品质性状的全基因组选择研究张爱民中国科学院遗传与发育生物学研究所中国科学院优秀通过2014CB138200玉米产量和品质性状全基因组选择育种的基础研究王国英中国农业科学院作物科学研究所农业农村部良好通过2014CB138300食用菌产量和品质形成的分子机理及调控张金霞中国农业科学院农业资农业农村部良好通过源与农业区划研究所2014CB138400农作物重要病毒病昆虫传播与致害的生物学基础李毅北京大学教育部优秀通过2014CB138500猪繁殖力的生理学及相关遗传调控机理崔胜中国农业大学教育部良好通过2014CB138600养殖鱼类蛋白质高效利用的调控机制麦康森中国海洋大学山东省科学技术厅、教育部优秀通过2014CB138700重要牧草、乡土草抗逆优质高产的生物学基础南志标兰州大学教育部良好通过2014CB138800天然草原生产力的调控机制与途径侯向阳中国农业科学院草原研究所农业农村部良好通过2014CB147200芽麦对品质的影响及抗穗发芽小麦材料全基因组选育王际睿四川农业大学四川省科学技术厅良好通过2014CB147300玉米叶型建成分子机理研究贺岩中国农业大学教育部良好通过2014CB238900煤中有害元素分布富集机理及环境污染防治代世峰中国矿业大学(北京)教育部良好通过2014CB239000中国陆相致密油(页岩油)形成机理与富集规律邹才能中国石油集团科学技术研究院中国石油天然气集团公司良好通过2014CB239100中国东部古近系陆相页岩油富集机理与分布规律黎茂稳中国石油化工股份有限公司石油勘探开发研究院中国石油化工集团公司良好通过2014CB239200超临界二氧化碳强化页岩气高效开发基础李晓红武汉大学教育部优秀通过2014CB239300基于半导体人工光合成的二氧化碳能源化研究叶金花天津大学教育部、天津市科学技术委员会良好通过2014CB239400人工光合成太阳能燃料的基础李灿中国科学院大连化学物理研究所中国科学院优秀通过2014CB239500大容量直流电缆输电和管道输电关键基础研究何金良清华大学教育部良好通过2014CB239600微型能源动力系统的科学问题赵黛青中国科学院广州能源研究所中国科学院良好通过2014CB239700基于超级电容器的大容量储能体系及其应马紫峰上海交通大学上海市科学技术委员优秀通过用会、教育部2014CB247400柔性直流输电换流器安全运行裕度的基础研究李武华浙江大学教育部、浙江省科学技术厅优秀通过2014CB247500燃气轮机高效清洁柔和燃烧机理及燃烧室基础研究张哲巅中国科学院工程热物理研究所中国科学院良好通过2014CB249200可再生能源与天然气融合的分布式能源的网络系统基础研究甘中学新奥科技发展有限公司河北省科学技术厅良好通过2014CB339800新型太赫兹源、接收器和其它关键功能器件及其应用陈健南京大学教育部良好通过2014CB339900无线接入网高能效微波集成器件理论及实现机理刘元安北京邮电大学教育部良好通过2014CB340000基于光子轨道角动量(OAM)的新型通信体制研究余思远中山大学教育部良好通过2014CB340100多维复用光纤通信基础研究李桂芳天津大学教育部、天津市科学技术委员会良好通过2014CB340200临近空间高速飞行器等离子鞘套信息传输吕跃广西安电子科技大学教育部优秀通过理论2014CB340300网络信息空间大数据计算理论怀进鹏北京航空航天大学工业和信息化部优秀通过2014CB340400网络大数据计算的基础理论及其应用研究华云生中国科学院计算技术研究所中国科学院良好通过2014CB340500面向三元空间的互联网中文信息处理理论与方法孙茂松清华大学教育部良好通过2014CB340600云计算安全基础理论与方法研究金海华中科技大学教育部良好通过2014CB340700安全攸关软件系统的构造与质量保障方法研究张健中国科学院软件研究所中国科学院良好通过2014CB347600大规模异质数据分析、挖掘与管理唐金辉南京理工大学江苏省科学技术厅良好通过2014CB347700基于情境的安全攸关软件的构造方法与运行机理研究熊英飞北京大学教育部良好通过2014CB347800软件定义的云数据中心网络基础理论与关键技术李丹清华大学教育部良好通过2014CB349300基于DPS的感知灵敏度拓展理论与关键技邓中翰北京中星微电子有限公北京市科学技术委员会良好通过术司2014CB440800新疆北部古弧盆体系成矿机理陈华勇新疆自然资源与生态环境研究中心新疆维吾尔自治区科学技术厅良好通过2014CB440900华南大规模低温成矿作用胡瑞忠中国科学院地球化学研究所中国科学院、自然资源部优秀通过2014CB441000东南丘陵区红壤酸化过程与调控原理沈仁芳中国科学院南京土壤研究所中国科学院良好通过2014CB441100土壤复合有机污染特征、界面行为及修复技术原理朱利中浙江大学教育部、浙江省科学技术厅优秀通过2014CB441200大气污染物的理化特征及其与气候系统相互作用廖宏中国科学院大气物理研究所中国科学院良好通过2014CB441300年代际尺度上全球和中国大气成分与气候的变化及其相互作用王斌清华大学教育部优秀通过2014CB441400雷电重大灾害天气系统的动力-微物理-电过程和成灾机理郄秀书中国科学院大气物理研究所中国科学院、中国气象局良好通过2014CB441500南海陆坡生态系统动力学与生物资源的可周朦上海交通大学上海市科学技术委员良好通过持续利用会、教育部2014CB447900城市边界层理化结构与成霾交互作用机制研究孙业乐中国科学院大气物理研究所中国科学院良好通过2014CB448000西天山石炭-二叠纪构造演化与浅成低温成矿系统蔡克大中国科学院新疆生态与地理研究所新疆维吾尔自治区科学技术厅、中国科学院良好通过2014CB541600出血性脑卒中发生发展机制及其防治的基础研究冯华中国人民解放军第三军医大学中央军委后勤保障部卫生局、重庆市科学技术委员会良好通过2014CB541700单基因遗传性聋病的分子机制研究管敏鑫浙江大学教育部、浙江省科学技术厅良好通过2014CB541800强直性脊柱炎发生机制及控制策略研究徐沪济中国人民解放军第二军医大学上海市科学技术委员会优秀通过2014CB541900系统性红斑狼疮的发病机理解析和诊治新策略研究沈南上海交通大学上海市科学技术委员会良好通过2014CB542000肿瘤异质性演化机制与个体治疗策略的生吴仲义中国科学院北京基因组中国科学院良好通过物学研究研究所2014CB542100肿瘤免疫逃逸新机制和免疫治疗新途径的基础与应用研究王青青浙江大学浙江省科学技术厅优秀通过2014CB542200周围神经损伤及修复后神经再生与中枢神经重塑的机制研究姜保国北京大学教育部良好通过2014CB542300循环miRNA生物学功能及临床应用张辰宇南京大学教育部优秀通过2014CB542400基于内皮细胞应激的糖尿病继发血管病变的早期关键机理与干预策略的研究陈丰原中南大学教育部良好通过2014CB542500慢性病毒感染的体液应答机制及功能重塑祁海清华大学教育部良好通过2014CB542600动物病毒-宿主相互作用机制的研究蒋争凡北京大学教育部良好通过2014CB542700猪繁殖与呼吸综合征病毒与宿主相互作用调控病毒复制及宿主免疫应答的机制杨汉春中国农业大学教育部良好通过2014CB542800新发、再发传染病病原体的结构研究饶子和南开大学教育部、天津市科学技术委员会优秀通过2014CB542900中医证候临床辨证的基础研究胡镜清中国中医科学院国家中医药管理局良好通过2014CB543000“上火”的机理与防治研究范永升浙江中医药大学国家中医药管理局、浙江省科学技术厅良好通过2014CB543100腧穴配伍方案优选及效应影响因素研究王之虹长春中医药大学吉林省科学技术厅、国家中医药管理局优秀通过2014CB543200腧穴配伍效应规律及神经生物学机制研究熊利泽中国人民解放军第四军医大学中央军委后勤保障部卫生局、国家中医药管理局良好通过2014CB548100表观遗传调控的中央杏仁核GABA神经环路与慢性神经痛张智中国科学技术大学中国科学院良好通过2014CB548200外周神经损伤引起中枢可塑性改变的能量代谢机制李相尧西安交通大学教育部良好通过2014CB643300海洋工程装备材料腐蚀与防护关键技术基础研究李晓刚中国科学院宁波材料技术与工程研宁波市科学技术局、中国科学院良好通过究所2014CB643400战略有色金属非传统资源清洁高效提取的基础研究冯其明中南大学教育部良好通过2014CB643500高效有机/聚合物太阳电池材料与器件研究黄飞华南理工大学教育部优秀通过2014CB643600高性能聚酰亚胺薄膜和纤维材料制备中的结构与性能调控杨士勇中国科学院化学研究所中国科学院良好通过2014CB643700资源节约型高性能稀土永磁材料设计和可控制备沈保根中国科学院物理研究所中国科学院优秀通过2014CB643800新型高性能稀土发光材料的科学基础及应用张洪杰中国科学院长春应用化学研究所中国科学院良好通过2014CB6439002.8-4.0微米室温高性能半导体激光器材料和器件制备研究王庶民中国科学院上海微系统与信息技术研究所中国科学院、上海市科学技术委员会良好通过2014CB644000集成高通量实验与计算的钛合金快速设计杜勇中南大学湖南省科学技术厅、教育部良好通过2014CB648300可印刷塑料电子材料及其大面积柔性器件赖文勇南京邮电大学江苏省科学技术厅优秀通过相关基础研究2014CB648400新型紫外/深紫外硼酸盐非线性光学材料的设计与制备研究杨志华中国科学院新疆理化技术研究所新疆维吾尔自治区科学技术厅良好通过2014CB744100航天飞行器跨流域空气动力学与飞行控制关键基础问题研究李志辉中国空气动力研究与发展中心中央军委装备发展部、四川省科学技术厅良好通过2014CB744200深空探测高精度天文测角测速组合自主导航基础研究张伟上海卫星工程研究所中国航天科技集团公司、上海市科学技术委员会优秀通过2014CB744300油页岩高效油气炼制与过程节能科学基础许光文西南化工研究设计院有限公司四川省科学技术厅良好通过2014CB744500基于影像实时动态多元分子分型的乳腺癌精准诊疗关键技术研究卢光明中国人民解放军南京军区南京总医院江苏省科学技术厅、中央军委后勤保障部卫生局良好通过2014CB744600基于生物、心理多模态信息的潜在抑郁风险预警理论与生物传胡斌兰州大学教育部良好通过感关键技术研究2014CB744700黄土重大灾害及灾害链的发生、演化机制与防控理论彭建兵长安大学自然资源部、陕西省科学技术厅良好通过2014CB744800大型客机减阻机理和方法研究吴光辉中国商用飞机有限责任公司上海飞机设计研究院中国商用飞机有限责任公司、上海市科学技术委员会良好通过2014CB744900面向大型飞机综合化航空电子系统安全性基础问题研究王国庆中国航空无线电电子研究所中国航空工业集团公司良好通过2014CB745000南海关键岛屿周边多尺度海洋动力过程研究田纪伟中国海洋大学教育部良好通过2014CB745100微生物多细胞体系的设计与合成元英进天津大学教育部、天津市科学技术委员会优秀通过2014CB745200合成生物器件干预膀胱癌的基础研究蔡志明深圳大学深圳市科技创新委员会良好通过2014CB748500废水中污染物强化传递分离的关键问题白志山华东理工大学上海市科学技术委员会良好通过2014CB748600视网膜多模态医学影像处理与分析及其应陈新建苏州大学江苏省科学技术厅优秀通过用基础研究2014CB845300不确定信息下多体导航与控制的系统理论和数学基础孙振东中国科学院数学与系统科学研究院中国科学院良好通过2014CB845400高温高密核物质形态研究马余刚中国科学院上海应用物理研究所中国科学院、上海市科学技术委员会优秀通过2014CB845500强流高功率离子加速器物理及技术先导研究赵红卫中国科学院近代物理研究所中国科学院良好通过2014CB845600团簇多级结构的构筑与功能调控谢素原厦门大学教育部、厦门市科学技术局良好通过2014CB845700基于LAMOST大科学装置的银河系研究及多波段天体证认刘晓为北京大学教育部优秀通过2014CB845800伽玛射线暴与相关前沿物理研究戴子高南京大学教育部良好通过2014CB845900核幔耦合作用与亚年代至世纪尺度地球自转及磁场变化关系研究倪四道中国科学院测量与地球物理研究所中国科学院良好通过2014CB846000光电子调控矿物与微鲁安北京大学教育部良好通过生物协同作用机制及其环境效应研究怀2014CB846100学习行为发生、发展及异常的认知神经机制研究李武北京师范大学教育部良好通过2014CB848700自旋及其复合系统的量子操纵与相干集成研究赵楠北京计算科学研究中心中国工程物理研究院良好通过2014CB848800分子铁电多功能存储材料的前沿交叉研究付大伟东南大学江苏省科学技术厅良好通过2014CB848900光催化体系表界面电子态的耦合与演化规律研究江俊中国科学技术大学中国科学院优秀通过蛋白质研究等6个重大科学研究计划2014CB910100先天免疫相关蛋白质复合物结构与功能的研究柴继杰清华大学教育部良好通过2014CB910200神经生长与信息传递重要蛋白质复合体的结构生物学研究张明杰深圳北京大学香港科技大学医学中心深圳市科技创新委员会优秀通过2014CB910300重要新型膜离子通道门控和调节机制的结构基础徐天乐上海交通大学教育部、上海市科学技术委员会良好通过2014CB910400蛋白质复合体和膜蛋白结构生物学中的新技术刘志杰中国科学院生物物理研究所中国科学院优秀通过和新方法研究2014CB910500mTOR调控网络在衰老及其相关代谢疾病发生进程中的作用及分子机制刘峰中南大学教育部良好通过2014CB910600肿瘤代谢异常的关键蛋白质作用机制及其分子调控网络高平中国科学技术大学中国科学院良好通过2014CB910700复杂生物大分子复合体的低温电镜高分辨三维重构及功能研究孙飞中国科学院生物物理研究所中国科学院优秀通过2014CB910800NOD样受体的免疫生物学及其相关疾病机制研究周荣斌中国科学技术大学中国科学院良好通过2014CB920900新型低维体系量子输运和拓扑态的研究杜瑞瑞北京大学教育部良好通过2014CB921000过渡金属氧化物异质结在多场调控下的新奇物性及器件研究金奎娟中国科学院物理研究所中国科学院良好通过2014CB921100低维磁性耦合体系的新物性及多场调控徐永兵南京大学教育部、江苏省科学技术厅良好通过2014CB921200多重量子相干器件制备、表征及外场调控许祝安浙江大学教育部、浙江省科学技术厅优秀通过2014CB921300新型高品质微腔中的光子与电子态耦合程亚中国科学院上海光学精密机中国科学院、上海市科学技优秀通过械研究所术委员会2014CB921400固体量子计算的器件物理基础刘玉玺清华大学教育部良好通过2014CB921500高轨道d电子体系的高压研制与强自旋-轨道耦合研究龙有文中国科学院物理研究所中国科学院良好通过2014CB921600能谷-自旋耦合量子态的光电作用机理和操控研究吴施伟复旦大学教育部、上海市科学技术委员会优秀通过2014CB931700液相激光熔蚀及在若干光电纳米材料制备中的关键科学问题研究杨国伟中山大学教育部良好通过2014CB931800密闭舱室环境安全保障纳米复合材料唐智勇国家纳米科学中心中国科学院优秀通过2014CB931900核酸药物高效纳米载药系统的研究申有青浙江大学教育部、浙江省科学技术厅良好通过2014CB932000典型人工纳米材料的水环境过程、生物效应及其调控研究刘思金中国科学院生态环境研究中心中国科学院良好通过2014CB932100纳米插层材料功能组装与盐湖镁资源有效利用段雪北京化工大学教育部优秀通过2014CB932200利用纳米材料与技术提高农药有效性与安全性的基础研究崔海信中国农科院农业环境与可持续发展所农业农村部良好通过2014CB932300二次锂空气电池高效能量转换与储存纳米材料的设计与调控周豪慎南京大学江苏省科学技术厅、教育部良好通过2014CB932400微纳超结构碳材料的设计制备及高效能量转换与存储研究康飞宇清华大学深圳研究生院深圳市科技创新委员会、教育部良好通过2014CB932500二维原子晶体界面科学与器件基础彭海琳北京大学教育部优秀通过2014CB932600有机纳米聚集体界面分子取向调控及光电器件应用基础研究唐建新苏州大学江苏省科学技术厅良好通过2014CB932700合成气转化制备优质液体燃料的高效金属纳米催化剂研究曾杰中国科学技术大学中国科学院良好通过2014CB942800神经元迁移、形态发生和微环路形成的调控机制张永清中国科学院遗传与发育生物学研究所中国科学院优秀通过2014CB942900脊柱发育与稳态维持的分子调控机制研究陈林中国人民解放军第三军医大学中央军委后勤保障部卫生局、重庆市科学技术委员会良好通过2014CB943000听觉微环路障碍形成的遗传和分子机制王秋菊中国人民解放军总医院中央军委后勤保障部卫生局良好通过2014CB943100精子发生与成熟的表观陈德桂中国科学院上中国科学院、良好通过遗传调控海生命科学研究院上海市科学技术委员会2014CB943200决定卵母细胞质量的分子机制苏友强南京医科大学卫生健康委、江苏省科学技术厅良好通过2014CB943300环境内分泌干扰物对女性生殖功能的影响及分子机制张军上海交通大学上海市科学技术委员会、教育部良好通过2014CB943400植物胚胎及种子发育的机理研究刘春明中国科学院植物研究所中国科学院优秀通过2014CB943500叶片发育极性建成的调控网络研究焦雨铃中国科学院遗传与发育生物学研究所中国科学院优秀通过2014CB943600淋巴细胞发育中的基因转录后调节网络研究常兴中国科学院上海生命科学研究院中国科学院、上海市科学技术委员会良好通过2014CB953700大气物质沉降对海洋氮循环与初级生产过程的影响及其气候效应高会旺中国海洋大学教育部、山东省科学技术厅优秀通过2014CB953800我国活性氮源及其对空气质量与气候变化的影响机理研究蔡祖聪南京师范大学江苏省科学技术厅良好通过2014CB953900全球变化背景下南海及周边地区春夏气候变异杨崧中山大学教育部良好通过。
鱼类基因组操作与定向育种.

中国科学: 生命科学2014年第44卷第12期: 1253 ~ 1261 SCIENTIA SINICA Vitae 引用格式: 叶鼎, 朱作言, 孙永华. 鱼类基因组操作与定向育种. 中国科学: 生命科学, 2014, 44: 1253–1261英文版见: Ye D, Zhu Z Y, Sun Y H. Fish genome manipulation and directional breeding. Sci China Life Sci, 2015, 58, in press 《中国科学》杂志社SCIENCE CHINA PRESS鱼类生物学和生物技术专辑评述鱼类基因组操作与定向育种叶鼎, 朱作言, 孙永华*中国科学院水生生物研究所, 淡水生态与生物技术国家重点实验室, 武汉 430072* 联系人, E-mail: yhsun@收稿日期: 2014-09-15; 接受日期: 2014-10-29国家重点基础研究发展计划(批准号: 2010CB126306, 2012CB944504)、国家自然科学基金(批准号: 31222052)、中国科学院知识创新工程重要方向项目(批准号: KSCX2-EW-N-004-4)和淡水生态与生物技术国家重点实验室自主研究项目(批准号: 2011FBZ23)资助摘要水产养殖已成为全球范围内发展最快的农业产业之一, 可持续发展水产养殖的关键在于培育具有优良性状的养殖品种. 基因组操作技术为快速、定向的鱼类遗传育种提供了一条重要的可行性途径. 本文回顾了基于经典基因组操作技术的鱼类育种方法学历史, 如多倍体育种及细胞核移植等. 然后重点介绍并展望了基于转基因技术及新近发展的基因组编辑技术的鱼类定向育种方法. 后两种技术的发展和应用将会为未来的鱼类种业带来更加高效和更具预见性的育种新方法. 关键词鱼类定向育种多倍体育种细胞核移植转基因鱼基因组编辑鱼类蛋白是人类最为重要的蛋白质来源之一. 由于过度捕捞导致渔业资源衰竭, 水产养殖已成为全球范围内最受关注和发展最快的农业产业之一[1]. 可持续发展水产养殖的关键在于培育具有优良性状的养殖品种. 现阶段, 鱼类养殖面临着近亲繁殖所带来的种质退化、鱼病频发、产量和品质下降等问题[2]. 因此, 筛选和培育高产、抗病、优质的养殖品种是可持续发展渔业的重中之重.在过去几十年间, 种内杂交[3]和种间杂交[4]等传统育种方法给人们提供了大量的优质鱼类产品. 然而, 杂交育种通常需要多代的循环选育才能造就具有某一优良性状且不表现出负面效应的新品种. 另外,利用这些方法, 也无法窥见这些优良性状背后的遗传机制, 使得选育品种的目标性状往往具有不可预见性. 因此, 亟需开发高效并可预测的育种方法来培育高产优质的鱼类新品种. 本文首先回顾基于经典基因组操作技术的鱼类育种方法学历史, 如多倍体育种[5]及细胞核移植[6]等. 然后, 重点介绍基于转基因技术及新近发展的基因组编辑技术的鱼类定向育种方法. 后两种技术的发展将会给未来的鱼类种业带来更加高效和更具预见性的育种新方法, 而基于经典和新兴基因组操作技术相结合的综合育种技术将极有可能成为未来鱼类育种的主导技术.1 多倍体育种多倍体育种通常通过倍性操作来实现. 倍性操作是一种通过人工干预致使染色体加倍的方法, 它可以说是基因组操作中最为经典的方法[5]. 在一些鱼类物种中, 雌、雄性或不育个体往往表现出不同的生长速率, 因此如何高效获得单性群体或不育群体往往是鱼类育种学家追求的目标. 在所有有关鱼类性别控制或育性控制的方法中, 倍性操作是使用最早也是目前为止最为有效的方法之一. 此外, 这一方法叶鼎等: 鱼类基因组操作与定向育种1254也被广泛用于纯系的快速获得, 加之与其他育种方法相结合, 将会大大缩减育种周期.鱼类多倍体育种源于对雌核发育的研究. 在鱼类中, 第二次减数分裂在排卵或受精后很短的时间内完成. 研究者发现, 这一过程能够被冷休克所抑制, 从而使卵细胞的染色体加倍[7,8]. 后续的实践表明, 热休克和静水压相结合的方法亦可获得类似的效果, 且更为简洁高效[9,10]. 通常而言, 鱼类人工雌核发育的实现是首先利用灭活的精子与正常成熟的卵子受精, 然后通过冷热激或静水压处理该受精卵发育而来[11,12]. 反之, 人工雄核发育是利用灭活的卵子与正常成熟的精子受精, 然后通过冷热激或静水压处理该受精卵发育而来. 雌核发育和雄核发育已在各种经济鱼类中得到广泛运用, 并获得双单倍体(double haploid)[10].双单倍体的获得能够被用来维持单一性别群体. 在XX-XY 性别决定机制的物种中, 雌核发育将会获得全雌后代; 而在雄核发育的后代中, 将会得到XX 和YY 2种基因型. 在ZW-ZZ 性别决定机制的物种中, 情况则相反. 由于辐射、温度、压力等处理对胚胎造成损伤从而导致胚胎具有较高的死亡率, 因此直接利用雌核发育或者雄核发育技术仅能生产少量的单性后代, 并不适宜于大规模的应用[10]. 然而, 值得强调的是, 多倍体育种常常会和一些经典的育种方法—如性逆转法相结合从而实现控制性别的目的. 全雄黄颡鱼(Pelteobagrus fulvidraco )研制则充分利用了这一策略[13]. 首先, 通过激素诱导性逆转的方法获得生理雌性的XY 型黄颡鱼, 再通过雌核发育获得YY 型超雄鱼. 当该超雄鱼和普通的XX 型雌鱼交配即可得到全雄后代. 另外, 利用激素诱导YY 型超雄鱼可获得生理雌性的YY 型雌鱼, 利用它们与正常YY 超雄鱼的交配可以维持一个有相当规模的超雄 亲本.与很多家养动物一样, 不育的鱼类品系往往比可育品系具有更快的生长速度. 因此, 三倍体不育鱼的育种在不同鱼类物种中获得广泛的研究和应用. 早期研究者们利用冷或热刺激正常的受精卵来获得三倍体不育鱼. 然而, 这种方法成功率较低并常常伴随着较高的死亡率. 相比之下, 利用四倍体鱼和二倍体鱼杂交来获得三倍体鱼则是一个更为高效的办 法[14]. 因为大多数的鱼都是二倍体, 所以如何获得具有繁殖能力的四倍体群体是该策略中最为关键的一步.一个最为成功的案例是利用红鲫(Carassius auratus )和鲤鱼(Cyprinus carpio )杂交直到F 3代, 通过筛选获得两性可育的异源四倍体鲫鲤[15,16]. 同一课题组也报道了利用红鲫和团头鲂(Megalobrama amblycephala )杂交成功获得了两性可育的异源四倍体鲫鲂[17,18]. 虽然多倍体育种在鱼类育种历史上存在已久, 并且利用此方法已经获得了多个优良品种, 但是利用这种方法建立四倍体品系并不具有普遍意义. 这主要是因为只能在为数不多的鱼类物种中才能通过杂交手段获得两性可育的四倍体群体. 此外, 三倍体不育系的最主要优良性状表现为快速生长, 因而这一方法并不适用于培育具有其他目标性状的品系. 当然, 这一方法也不适用于一些雌、雄性或不育个体在生长上没有显著差别的鱼类品种.2 细胞核移植细胞核移植被认为是最为彻底的一种基因组操作. 这一技术将一整套细胞核基因组移植到另一个去核的卵细胞中, 从而构建一个重构卵. 如果移植的细胞核和卵细胞来源于同一物种, 这种细胞核移植被称作“同种核移植”. 同种核移植已被广泛用于研究细胞核的可塑性、细胞核的再程序化以及利用体细胞核获得再程序化的干细胞等相关研究[19]. 虽然自20世纪50年代以来, 细胞核移植(动物克隆)技术已在多个物种中获得了较为广泛的研究, 利用这一技术也成功地获得了干细胞来源的克隆动物[6], 但是直到首例体细胞克隆哺乳动物“多莉”羊的诞生才掀起了另一波研究的热潮[20]. 在鱼类中, 细胞核移植技术最早由童第周等人[21]在金鱼(Carassius auratus )和鳑鲏(Rhodeus sinensis )中建立. 其实, 早在1985年, 陈宏溪等人[22,23]报道了第一例来自于短期培养体细胞的克隆鱼, 但是这一研究因为最初以中文发表而并不为国际学术界所知. 2002年, Lee 等人[24]报道了利用长期培养的斑马鱼(Danio rerio )细胞获得克隆斑马鱼, 从而点亮了利用基因操作后的鱼类培养细胞进行个体重建的希望之光. 有意思的是, 在青鳉鱼(Oryzias latipes )中, “半克隆”的技术也能获得成功—研究者利用单倍体胚胎干细胞进行细胞核移植, 从而获得了单倍体克隆鱼[25]. 然而, 不得不指出, 体细胞克隆的成功率非常低, 而且克隆子代往往出现各种发育缺陷. 因而, 必须解决以上问题, 才能有效中国科学: 生命科学 2014年 第44卷 第12期1255地将克隆技术应用于鱼类遗传育种.如果用于克隆的卵细胞和供体细胞来源于2个不同的物种, 则被称作“异种核移植”. 由于异种核移植可以将来源于不同物种的细胞核和卵细胞质因子进行组合, 因此认为对鱼类育种具有较大的价值[26]. 异种核移植最早在两栖类的2个属—Rana 或Xenopus 内的不同物种间进行[27,28]. 在这些实验中, 研究者无一例外地无法获得成功的克隆子代. 可能原因是供体细胞核的不完全再程序化或用于克隆的物种间的核质不相容性. 在哺乳动物的近缘物种间, 已有若干的濒危动物获得成功克隆[29]. 这些实验同时显示, 克隆动物在各个方面与细胞核供体完全一致, 显示细胞核基因组对性状的决定性作用. 然而, 在具有不同表型的鱼类不同物种间开展的细胞核移植研究给出了非常不一样的结果.作为脊椎动物中较为低等的类群, 鱼类中的异种细胞核移植似乎可以获得更广泛的成功. 为了获得具有优良经济性状的“核质杂交鱼”, 童第周课题组[26]利用跨属的鲤鱼细胞核和鲫鱼去核卵进行细胞核移植试验并获得成功, 以及将鲫鱼细胞核移入鲤鱼去核卵也能获得成功[30]. 在童第周等人开展的实验中, 以及本课题组近期将转基因鲤鱼细胞核移入金鱼去核卵所获得的属间克隆鱼实验中[31], 均可以观察到克隆鱼的脊椎骨数目与受体鱼一致, 而不同于供体鱼. 这表明, 鱼类卵细胞质不仅能驱动来自于相对远缘物种的供体细胞核的发育, 并且能够在一定程度上影响克隆鱼的发育和性状决定. 鱼类的异种核移植甚至在不同科间—如金鱼(Carassius auratus , family Cyprinidae, order Cypriniformes)和泥鳅(Paramisgurnus dabryanus , family Cobitidae, order Cypriniformes)间, 以及不同目间—如罗非鱼(Oreochromis nilotica , order Perciformes)和金鱼间展开[32,33]. 但是, 所有这些科间或目间的细胞核移植均无法获得成功. 这不得不重新检视这些已获得成功的鱼类异种克隆, 结果发现, 它们只能在那些原本就能够进行人工杂交的物种间获得成功. 同样, 异种克隆的效率依然低下. 因此, 很难想象利用异种克隆技术能够将远缘鱼类物种的优良性状导入到经济鱼类品种中, 但其在珍稀或濒危鱼类的保护和育种中有着重要的应用前景.由于核移植的技术难度较大, 研究者们试图去寻找一种替代方法—细胞融合, 从而可以在一次实验中获得大量的重构卵. 这种方法已经成功的用于获得鲤鱼细胞核与银鲫(Carassius auratus, genus Carassius )卵细胞质的核质杂交鱼[34]. 这种杂交鱼显示出与核供体鲤鱼相似的表型. 此外, 另一项有意义的研究利用抗草鱼(Ctenopharyngodon idellus )出血病病毒的肝细胞与未受精卵融合来创建品系[35]. 然而, 对该品系的进一步的抗病性分析却未见报道. 同核移植相比, 通过细胞融合来获得幼鱼的成功率同样极低, 这在很大程度上限制了该方法在育种研究和品系建立中的应用.3 转基因育种随着功能基因组学的飞速发展, 从植物到动物, 从非脊椎动物到脊椎动物, 成千上万种基因的功能获得阐释并表现出物种间的功能保守性. 理论上, 通过转基因技术, 可以在最短时间内使得目标鱼类获得某一(些)特定基因所关联的性状, 并且这些性状是稳定和可遗传的.显微注射是最广为使用的生产转基因鱼的方 法[36]. 其他的基因转移方法在鱼类转基因研究中也被广泛应用, 包括电转法[37~39]、精子介导法[40,41]、电脉冲精子介导法[42,43]、逆转录病毒法[44~46]和脂质介导法[47]等. 世界上首例转基因鱼在中国诞生—研究者们利用小鼠金属硫蛋白Ⅰ型基因的启动子驱动人生长激素基因对金鱼进行转基因实验[36]. 后来, 生长激素转基因在各种鱼类中被加以广泛研究. 例如, 各种生长激素基因被转移到泥鳅[48]、鲤鱼[49~53]、大西洋鲑鱼(Salmo salar )[54]、鲶鱼(Silurus asotus )[39]、罗非鱼[55,56]等经济鱼类中.除了通过转入生长激素获得更高的生长速率外, 其他的基因也被成功导入到鱼中以期获得抗逆形状. 例如, 抗冻蛋白转基因被用来增强鱼类的抗冻能 力[54,57]; 溶菌酶基因被导入到大西洋鲑鱼中以使其获得抗病特性[58,59]; 天蚕(Hyalophora cecropia )抗菌肽B 基因转基因鲶鱼表现出一定的抗病特性[60]; 人乳铁蛋白基因被导入到草鱼用以增强其抗出血病能力[43]; 透明颤菌血红蛋白基因被用于抗低氧转基因鱼的研究[61].出于对潜在的转基因安全和生物伦理的考虑, 研究者们考虑过表达鱼类自身来源的基因, 而不是外源性的人生长激素基因, 这被称为“全鱼”转基因.叶鼎等: 鱼类基因组操作与定向育种1256基于该策略, 一种新的“全鱼”生长激素转基因载体被成功设计和应用, 它是由鲤鱼的 -actin 基因的启动子驱动草鱼生长激素基因表达的[50,53]; 另外一个例子是用大洋鳕鱼(Zoarces americanus )的抗冻蛋白启动子介导大马哈鱼(Oncorhynchus keta )生长激素的cDNA 表达[54]. 在这2例研究中, 转基因鱼均表现出较非转基因鱼显著的生长优势.除了天然编码基因可用于转基因之外, 在鱼类转基因研究中也提出了更高效的“分子设计”转基因的概念. 在斑马鱼中转入了一种人为改造的激活型的生长激素受体基因, 此基因的表达产物生长激素受体可形成不依赖于生长激素结合的二聚体状态, 从而持续地激活下游的信号通路. 持续激活型生长激素受体转基因鱼表现出较普通生长激素转基因鱼更高的生长速度[62]. 利用“分子设计”的方法来构建新型高效的目标基因, 将是转基因鱼研究值得重点发展的方向.“合成生物学”的概念也开始应用于鱼类转基因育种, 其目的是使鱼具有更高的营养价值. 长链多不饱和脂肪酸(polyunsaturated fatty acid, PUFA)对人类健康特别是神经系统的发育和健康具有重要功能[63], 而其来源极其有限. 研究者将鱼类来源的脂肪酸去饱和酶和延长酶导入斑马鱼受精卵, 用于生产富含n3及n6长链多不饱和脂肪酸的转基因鱼[64~66]. 然而, 由于鱼类缺乏将n6 PUFA 转化为n3 PUFA 的能力以及从头合成n6 PUFA 的能力, 这类转基因鱼体内的n3和n6 PUFA 水平依然依赖于饲料中n3或者n6 PUFA 的含量. 本课题组克隆了线虫(Caenorhabditis elegans )来源的∆12-去饱和酶基因(fat1)和∆15-去饱和酶基因(fat2), 并将其进行鱼类密码子优化, 通过以上2个基因向鱼类基因组的转移实现了长链多不饱和脂肪酸在鱼类物种中的从头合成. 对于这种转基因鱼, 即便投喂不含n3和n6 PUFA 的饲料, 依然可以在转基因鱼体内检测到高水平的n6和n3 PUFA 含量[67]. 转基因鱼还可以用于生物反应器的研究. 有研究者利用斑马鱼卵生产如人凝血因子Ⅶ[68]、黄体 素[69]以及类胰岛素样生长因子[70]等生物活性物质.如果希望获得集多种目标性状于一体的转基因鱼, 除了可以结合传统的杂交选育手段外, 也可以利用2A 短肽技术来实现. 2A 短肽可以用来有效地在同一细胞中表达被2A 序列分隔的多个基因. 该短肽被证明在包括鱼[71]、小鼠[72]、猪[73]等多个物种中有效.利用2A 短肽将可以使转基因鱼同时获得多个有价值的表型, 因而能够大大地缩短多目标性状转基因的育种周期.经遗传修饰过的动物, 可能存在入侵自然生态的潜在风险, 因此需要开展详尽的生态安全评估研究[74]. 除了广泛而深入地评价转基因鱼对其他物种的影响外[75], 一个可行的杜绝潜在生态安全的解决办法是生产不育的转基因鱼[76,77]. 在以上所有类型转基因鱼中, 有2类最接近于上市的品系. 一种是由AquaBonty 公司生产的转生长激素基因大西洋鲑, 此种转基因鱼已经提交至美国食品和药物管理局 等待上市批准(/AnimalVeterinary/ DevelopmentApprovalProcess/GeneticEngineering/Gen eticallyEngineeredAnimals/ucm280853.htm). 另一种是本课题组获得的三倍体转生长激素基因鲤鱼, 该鲤鱼表型出明显的快速生长优势且完全不育[76].4 基因组编辑近年来, 各种靶向核酸酶介导的基因组编辑技术在斑马鱼等鱼类物种中得到飞速发展, 如锌指核酸酶(zinc finger nucleases, ZFNs)[78]、转录激活因子样效应物核酸酶(transcription activator-like effector nucleases, TALEN)[79,80]和CRISPR(clustered regularly interspersed short palindromic repeats)/Cas9[81~83]技术等. 相对于ZFNs 和TALENs 而言, CRISPR/Cas9系统组分较为简单并可达到相似甚至更高的效率, 因而CRISPR/Cas9系统成为发展最快的基因组编辑技术并且广为所用. CRISPR/Cas9系统利用一个短的通常含有约20个碱基靶序列的向导RNA(gRNA)结合到它的DNA 互补靶点, 并介导Cas9核酸酶到此靶点切断靶序列从而造成双链断裂(double-strand breaks, DSBs). 这些DSBs 一般通过非同源末端连接方式(nonhomologous end-joining, NHEJ)或者通过同源介导的DNA 修复机制(homology-directed repair, HDR)进行修复. NHEJ 往往导致突变的产生, 而HDR 在外源DNA 修复模板的介导下可以实现精细的基因组编辑[84]. 目前, 将这一技术成功运用到育种研究的报道仅有少数几例. 凯尔特(Celtic) POLLED 基因位点是无角牛表现出无角性状的等位位点, 研究者们利用 TALEN 介导的精细基因组编辑技术将POLLED 位点引入到普通有角奶牛基因组中,意图使普通有角奶牛中国科学: 生命科学 2014年 第44卷 第12期1257获得无角性状[85]. 在鱼类中, 利用斑马鱼模型开展了针对GH(growth hormone)信号通路最主要的负调节因子SOCS(suppressor of cytokine signaling)家族基因socs2的敲除试验, 结果显示, socs2敲除的斑马鱼表现出GH 信号通路下游靶基因的激活且在胚胎发育早期表现出一定的快速生长效应[86].单链核苷酸被证明是一种有效的介导HDR 的修复模板, 它通过单链复性(single strand annealing, SSA)的机制来实现HDR. 通过共注射靶向核酸酶和单链DNA, 已成功地在斑马鱼和小鼠中导入了单核苷酸的改变[79,87]. 从科学的角度而言, 这种单核苷酸的改变当然可以等同于在自然界固有存在的单核苷酸多态性(single nucleotide polymorphism, SNP). 这一方法也能够被用于在特定位点引入诸如HA(hemagglutinin)标签, loxP 位点等短的DNA 片 段[79,82,88]. 然而, 主要由非特异性NHEJ 导致的脱靶效应常常发生. 新改进的Cas9通过RuvC 或者HNH 核酸酶结构域产生点突变, 使其仅能在一条链上产生断裂而不是像普通Cas9那样产生双链断裂[89~91]. 用这种突变的Cas9结合一对引导RNA 能够介导高效的插入或者缺失突变, 与普通Cas9相比, 可将非特异性最高降低1500倍, 从而大大降低脱靶效应[92,93].另外, TALEN 介导的同源重组(homologous recombination, HR)已经在斑马鱼中获得初步的成 功[94]. CRISPR/Cas9介导的同源重组也在线虫[95]、果蝇(Drosophila melanogaster)[96~98]、人类细胞[99,100]上获得成功. 虽然靶向核酸酶技术已经大大地增加了鱼类胚胎中同源重组的效率, 但是该效率还远达不到满意的程度. 为了提高同源重组的效率, 研究表明, 通过抑制一些参与NHEJ 基因的功能来抑制其效 率, 可以有效的提高同源重组的效率[101~105]. 很多研究表明, 同源介导的DNA 修复常发生在体细胞, 同源重组子代的筛选过程冗长并且耗费人力. 如果生殖细胞特异的同源重组在F 0中可以被快速地检测出来, 将大大节省筛选的工作量. 基于UAS/ Gal4和Cre/loxP 系统构建的原始生殖细胞(primordial germ cell, PGC)特异的操作工具将有可能提供上述基于PGC 检测的快速筛选平台[106].基因组编辑技术能够高效地并在相对较短的时间内特异地对多个基因进行精细编辑[107], 因而它非常有利于对现有的优良品系进行改良. 更重要的是, 利用单链DNA 退火介导的单碱基替换技术不涉及新DNA 元件的引入或大片段的缺失, 其最终存在形式可以完全等同于自然界广泛存在的单核苷酸多态性. 因此, 如果将这项技术结合基于PGC 检测的筛选平台将会使得鱼类和其他动物育种的预见性更强、效率更高.5 总结科学技术的发展常常会加速科学研究的进程, 变“不可能”为“可能”. 转基因鱼的商品化虽然遭遇到一定的困难, 但事实表明, 没有一例批准上市的转基因食品出现安全问题. 研究者们需要做的是在转基因食品上市之前做好科学而严谨的评估工作, 而不是盲目地反对转基因食品. 除此之外, 高速发展的基因组编辑技术为鱼类育种研究提供更为宝贵的工具. 把精细基因组编辑技术与传统的鱼类育种技术相结合能够更加高效、更加自如和更具目的性地改善某一(或某些)特定性状, 而不去干扰其他任何性状. 由于这一技术不会引入任何的外源基因片段而仅对内源基因组作最为精细的编辑, 这将成为未来鱼类育种技术发展的重要方向.参考文献1 Cressey D. Aquaculture: future fish. Nature, 2009, 458: 398–4002 桂建芳, 朱作言. 水产动物重要经济性状的分子基础及其遗传改良. 科学通报, 2012, 57: 1719–17293 Bakos J, Gorda S. Genetic-improvement of common carp strains using intraspecific hybridization. Aquaculture, 1995, 129: 183–1864 Bartley D M, Rana K, Immink A J. The use of inter-specific hybrids in aquaculture and fisheries. Rev Fish Biol Fisher, 2000, 10:325–3375 Wu C J, Ye Y Z, Chen R D. Genome manipulation in carp (Cyprinus carpio L.). Aquaculture, 1986, 54: 57–616 Zhu Z Y, Sun Y H. Embryonic and genetic manipulation in fish. Cell Res, 2000, 10: 17–277 Sajiro Makino Y O. Formation of the diploid egg nucleus due to suppression of the second maturation division, induced by refrigerationof fertilized eggs of the carp, Cyprinus carpio . Cytologia, 1943, 13: 55–60叶鼎等: 鱼类基因组操作与定向育种8Nagy A, Rajki K, Horvath L, et al. Investigation on carp, Cyprinus carpio L. gynogenesis. J Fish Biol, 1978, 13: 215–2249Streisinger G, Walker C, Dower N, et al. Production of clones of homozygous diploid zebra fish (Brachydanio rerio). Nature, 1981, 291: 293–29610Komen H, Thorgaard G H. Androgenesis, gynogenesis and the production of clones in fishes: a review. Aquaculture, 2007, 269: 150–173 11Chen S L, Tian Y S, Yang J F, et al. Artificial gynogenesis and sex determination in half-smooth tongue sole (Cynoglossus semilaevis).Mar Biotechnol, 2009, 11: 243–25112Chen S L, Ji X S, Shao C W, et al. Induction of mitogynogenetic diploids and identification of ww super-female using sex-specific ssr markers in half-smooth tongue sole (Cynoglossus semilaevis). Mar Biotechnol, 2012, 14: 120–12813Wang D, Mao H L, Chen H X, et al. Isolation of Y- and X-linked scar markers in yellow catfish and application in the production of all-male populations. Anim Genet, 2009, 40: 978–98114Horvath L, Orban L. Genome and gene manipulation in the common carp. Aquaculture, 1995, 129: 157–18115Liu S J, Liu Y, Zhou G J, et al. The formation of tetraploid stocks of red crucian carp X common carp hybrids as an effect of interspecific hybridization. Aquaculture, 2001, 192: 171–18616Guo X H, Liu S J, Liu Y. Evidence for recombination of mitochondrial DNA in triploid crucian carp. Genetics, 2006, 172: 1745–174917Liu S J, Qin Q B, Xiao J, et al. The formation of the polyploid hybrids from different subfamily fish crossings and its evolutionary significance. Genetics, 2007, 176: 1023–103418Qin Q B, He W G, Liu S J, et al. Analysis of 5s rDNA organization and variation in polyploid hybrids from crosses of different fish subfamilies. J Exp Zool Part B, 2010, 314: 403–41119Gurdon J B, Wilmut I. Nuclear transfer to eggs and oocytes. Cold Spring Harb Perspect Biol, 2011, 3, doi: 10.1101/cshperspect.a002659 20Wilmut I, Schnieke A E, McWhir J, et al. Viable offspring derived from fetal and adult mammalian cells. Nature, 1997, 385: 810–81321童第周, 吴尚懃, 叶毓芬, 等. 鱼类细胞核的移植. 科学通报, 1963, 7: 60–6122陈宏溪, 易咏兰, 陈敏容, 等. 鱼类培养细胞核发育潜能的研究. 水生生物学报, 1986, 10: 1–723Chen H, Yi Y, Chen M, et al. Studies on the developmental potentiality of cultured cell nuclei of fish. Int J Biol Sci, 2010, 6: 192–19824Lee K Y, Huang H, Ju B, et al. Cloned zebrafish by nuclear transfer from long-term-cultured cells. Nat Biotechnol, 2002, 20: 795–79925Yi M, Hong N, Hong Y. Generation of medaka fish haploid embryonic stem cells. Science, 2009, 326: 430–43326Tung T C, Tung Y Y F. Nuclear transplantation in teleosts. I. Hybrid fish from the nucleus of carp and the cytoplasm of crucian. Sci Sin B, 1980, 14: 1244–124527Moore J A. Serial back-transfers of nuclei in experiments involving two species of frogs. Dev Biol, 1960, 2: 535–55028Gurdon J B. The transplantation of nuclei between two species of xenopus. Dev Biol, 1962, 5: 68–8329Sun Y H, Zhu Z Y. Cross-species cloning: influence of cytoplasmic factors on development. J Physiol, 2014, 592: 2375–237930Yan S, Lu D, Du M, et al. Hybrid fish from the nucleus of crucian and the cytoplasm of carp. Sci Sin B, 1984, 27: 1029–103431Sun Y H, Chen S P, Wang Y P, et al. Cytoplasmic impact on cross-genus cloned fish derived from transgenic common carp (Cyprinus carpio) nuclei and goldfish (Carassius auratus) enucleated eggs. Biol Reprod, 2005, 72: 510–51532Yan S Y, Mao Z R, Yang H Y, et al. Further investigation on nuclear transplantation in different orders of teleost: the combination of the nucleus of tilapia (Oreochromis nilotica) and the cytoplasm of loach (Paramisgurnus dabryanus). Int J Dev Biol, 1991, 35: 429–43533Yan S Y, Tu M, Yang H Y, et al. Developmental incompatibility between cell nucleus and cytoplasm as revealed by nuclear transplantation experiments in teleost of different families and orders. Int J Dev Biol, 1990, 34: 255–26634易咏兰, 刘沛霖, 刘汉勤, 等. 鱼类囊胚细胞和卵的电融合. 水生生物学报, 1988, 12: 189–19235余来宁, 左文功, 方耀林, 等. 用电融合结合继代移核方法构建草鱼抗病体细胞工程鱼. 水产学报, 1996, 20: 314–31836Zhu Z Y, Li G H, He L, et al. Novel gene transfer into the fertilized eggs of goldfish (Carassius auratus L.1758). J Appl Ichthyol, 1985, 1: 31–3437Inoue K, Yamashita S, Hata J, et al. Electroporation as a new technique for producing transgenic fish. Cell Differ Dev, 1990, 29: 123–128 38Xie Y, Liu D, Zou J, et al. Gene transfer via electroporation in fish. Aquaculture, 1993, 111: 207–21339Powers D A, Hereford L, Cole T, et al. Electroporation: a method for transferring genes into the gametes of zebrafish (Brachydanio rerio), channel catfish (Ictalurus punctatus), and common carp (Cyprinus carpio). Mol Mar Biol Biotechnol, 1992, 1: 301–30840Khoo H W, Ang L H, Lim H B, et al. Sperm cells as vectors for introducing foreign DNA into zebrafish. Aquaculture, 1992, 107: 1–1941Lavitrano M, Camaioni A, Fazio V M, et al. Sperm cells as vectors for introducing foreign DNA into eggs-genetic-transformation of mice.Cell, 1989, 57: 717–72342Tsai H J. Electroporated sperm mediation of a gene transfer system for finfish and shellfish. Mol Reprod Dev, 2000, 56: 281–28443Zhong J Y, Wang Y P, Zhu Z Y. Introduction of the human lactoferrin gene into grass carp (Ctenopharyngodon idellus) to increase resistance against gch virus. Aquaculture, 2002, 214: 93–1011258。
农业综合知识题(含答案)

农业综合知识(选择题 16 题,判断题 4 题,问答题 75 题,共 95 题)一、选择题1. 超级稻是指:( A ) A.产量特别高的水稻 B.米质特别好的水稻 C.抗性特别好的水稻 2 下列作物中,哪个不是我国主要粮食作物?( C ) A.玉米 B.大豆 C.高粱3.广东最大的粮食作物是:( A )A.水稻 B.玉米 C.马铃薯4.“绿色革命”发生在上世纪( B )A.30 年代B.50 年代C.70 年代5.水稻占广东粮食作物播种面积的( C ) A.60% B.70% C.80%6. 在有机食品生产过程中,是否可以使用化肥?( A )A.不可以B.可以C.无所谓7.根据我国的有关行业分类标准《国民经济行业分类》(GB/T4754-2002),畜牧业包括牲畜的饲养、猪的饲养、家禽的饲养、狩猎和捕捉动物、其他畜牧业,其中牲畜包括以下哪类动物:( A )A.牛的饲养B.猪的饲养C.鸡的饲养D.狗的饲养8.广东省地方畜禽品种资源十分丰富,列入《国家级畜禽遗传资源保护名录》的畜禽品种有 9 个,其中不包括以下哪一个品种:( D )A.蓝塘猪B.惠阳胡须鸡C.乌鬃鹅D.岭南黄鸡9. 广东省在配套系选育方面取得了很大成绩,先后培育出了 3 个猪配套系和1ⅠⅠ号配套系10. 根据国家有关规定,以下哪一种药物是国家明文禁止添加的违禁药物,不能作为饲料添加剂使用。
( B ) A.玉米 B.瘦肉精(盐酸克伦特罗)C.食盐D.维生素11. 以下哪一个国家是全世界养猪业最发达、猪肉出口最多的国家之一?( C )A.古巴B.朝鲜C.丹麦D.澳大利亚12. 农产品加工是把种植、养殖等的农副产品按其用途分别制成成品或半成品的生产过程。
针对原料的加工程度而言可分为(B)。
A.种植、初加工、深加工 B.养殖、初加工、深加工13. 我省粮食加工优势产业主要集中在大米加工、米面制品及烘焙食品、加工三大领域,优势产品包括(ABDFG)。
国家高技术研究发展计划(863计划)

一、指南说明海水养殖种子工程包括海水养殖优良种质创制和健康苗种繁育两个部份,是发展海水养殖业的重要物质基础,也是海洋农业技术领域国际竞争的焦点。
加快育种核心前沿技术的研究,哺育优良品种,实现名贵海水养殖生物苗种的规模繁育,对发展“高产、优质、抗逆、生态、安全”的海水养殖业,提升海洋农业科技核心创新能力和国际竞争力,促进我国向现代海洋农业强国转变具有重要意义。
本项目以“十五”期间863 计划、973 计划等在海水养殖种子工程方面取得的研究成果和建成的基地为基础,应用份子标记辅助育种技术,结合传统杂交和选育等技术,系统开展鱼类、虾蟹类、贝类、海参和藻类等的遗传改良研究,创制一批新种质材料;定位一批与重要生产性状相关的QTLs,构建份子育种技术体系;提升细胞工程育种技术、重要生产性状基因的发掘与应用技术、份子育种技术和鱼虾性别控制技术的水平,培植技术源头创新能力;开展繁殖调控技术、优质亲体和苗种哺育技术及优良品种的快速繁育技术等的研发。
此次发布的是现代农业技术领域“海水养殖种子工程”重大项目课题申请指南,拟支持16 个研究课题,包括:(1)重要海水养殖动物细胞工程育种技术;(2)重要生产性状关键基因的发掘与应用技术;(3) 鱼、虾等性别控制技术; (4)鲆鲽类高产、抗逆品种的哺育; (5)大黄鱼等优质、抗逆品种的哺育; (6) 虾、蟹高产、抗病品种的哺育;(7)鲍高产、抗逆品种的哺育;(8)扇贝高产、抗逆品种的哺育;(9) 珍珠贝和牡蛎优质、高产、抗逆品种的哺育;(10)蛤、蚶、蛏等高产、抗逆品种的哺育;(11)刺参、海胆等高产、抗逆品种的哺育;(12)海带、裙带菜等优质、高产、抗逆品种的哺育;(13)紫菜、江篱等优质、高产、抗逆品种的哺育;(14)名贵鱼类苗种规模繁育技术;(15)大珠母贝、江珧等苗种规模繁育技术;(16)羊栖菜等苗种规模繁育技术。
本重大项目安排国拨经费0.795 亿元。
二、指南内容课题1. 重要海水养殖动物细胞工程育种技术研究目标:建立海水动物的细胞系,哺育四倍体亲本群体。
水产动物育种学

水产动物育种学绪论1、水产动物育种:是指应用各种遗传学方法,改造水产动物的遗传结构,培育出适合人类养殖生产活动需要的品种的过程。
2、水产动物育种学:是研究水生生物育种理论和方法的科学,是水产养殖学的一个分支,也是研究野生种类驯化、优良物种引进、水产动物品质改良、繁育群体生产性能保护、杂种优势利用以及新品种培育的理论和实践的一门科学。
3、育种目标:对育种工作需要解决的主要问题的定性或定量的描述,也就是所要培育的新品种在一定自然、生产、经济及技术条件下养殖时,应具备的一系列优良性状的指标,是育种方案的基本内容之一。
4、制定育种目标的原则(论述):1.需要与可能:育种学家应该有丰富的想象力和科学的预见性.根据科学规律进行分析,把客观需要和现实需要的可能性结合起来构成现实的育种目标.制定育种目标时应考虑育种单位拥有的种质资源、技术力量、实验室及场地、设施、经费等因素是否具有实现育种目标的潜力.2.当前与长远:制定育种目标既要着眼于现实和近期内发展需要,同时也尽可能兼顾到长远发展需要。
在解决现目标时,不要把长远目标弃而不顾.在一个较长而复杂的目标内,制定出分阶段的育种目标。
3.目标性状和非目标性状:制定育种目标时应明确亟待改进的目标性状。
目标性状集中,相对选择压大,育种效率较高。
相反,如果目标性状分散,势必分散精力,延缓育种进度.目标性状一般不能超过2—3个,而且还要根据性状在育种中的难度和重要性,明确主要目标性状和次要目标性状,做到主次分明,协调改进。
切忌要求过宽齐头并进、主次不分、主次颠倒或过分忽视次要目标。
4.育种目标和组成性状的具体指标:育种目标尽可能简单明确,除了必须突出重点外,一定要把育种目标落实到具体性状上,而且尽可能提出数量化的可以检验的客观指标。
一个育种项目中可以包括少数几个性质相近的不同育种目标。
5、品种和品系:品种:是人类根据自身需要创造出的一种生产资料,一般是指经多代人工选育成的,具有遗传稳定,并有别于原种或同种内其他群体,具有优良的经济性状及其他表型性状的水生动物群体。
国家重点基础研究报告发展计划、国家重大科学研究报告计划重要支持方向

国家重点基础研究报告发展计划、国家重大科学研究报告计划重要支持方向个人资料整理仅限学习使用国家重点基础研究发展计划、国家重大科学研究计划2018年重要支持方向农业领域1.农作物优异亲本形成的遗传基础和优良基因资源合理组配与利用以主要粮食或油料作物优异亲本及其衍生的大面积推广品种系谱为基础,从表型组学、基因组学入手,系统分析优异亲本中的主要优良性状基因、基因组区段、组合方式及其遗传效应,筛选与创制对农业生产有重要推动作用的候选亲本,为新品种培育提供理论基础。
2.农作物产量、品质或抗性形成机制及其调控研究作物产量、品质、抗性等性状形成的生物学基础和遗传调控网络,建立整合生物学模型和品种分子设计的技术体系,培育高产、优质、环境友好的作物新品系;研究叶片衰老与作物产量、品质或抗性形成的关系及叶片衰老关键基因的调控作用,创新可用于遗传改良的转基因作物材料,为作物高产、优质分子育种提供理论支撑。
3.农作物抗逆、养分高效利用研究作物感受和应答环境胁迫或养分高效利用的遗传学基础,解读作物响应环境胁迫或养分高效利用的信号转导和分子调控网络,为农作物品种改良分子设计提供理论支持和基因工程育种模型。
4.农田地力培肥和作物多样性栽培的生态调控研究我国粮食主产区中低产田土壤地力要素形成机制、相互作用过程和改良措施,探索地力培肥和保育的科学途径,建立集约化条件下土壤地力持续提升标准,为实现作物高产稳产和环境友好的土肥水资源持续利用提供理论、方法;研究和评价典型区域作物多样性栽培增产技术对产品质量和农田生态环境<土壤、病虫等)的影响,为建立高产栽培技术规程和标准提供理论和技术指导。
5.畜禽重要经济性状形成机理与调控研究畜禽产品优良品质形成的分子遗传学基础和代谢调控网络,应用整合生理学和营养基因组学等理论与方法,设计和干预关键的网络调控通路,为改善畜禽品质提供基础理论和技术指导。
6.农作物重要病虫害防治和重要经济昆虫的基础研究建立和完善植物免疫研究的体系与技术平台,发展作物抗病机理研究的模式体系;研究基于基因沉默的害虫控制理论,阐明引发害虫基因沉默的RNA有效传导机制和害虫致死基因快速控制种群生长的途径,建立农作物抗病虫分子育种的新策略与新技术;研究家蚕基因功能注释和克隆重要突变基因,确立分子设计育种理论基础与技术体系。
鱼类基因组学及基因组育种技术

鱼类基因组学及基因组育种技术一、引言随着生物技术的飞速发展,鱼类基因组学及基因组育种技术已成为水产养殖领域的前沿研究领域。
了解这些技术对于改善鱼类生长性能、抗病性和环境适应性具有重要意义,有助于推动水产养殖业的可持续发展。
二、鱼类基因组学鱼类基因组学是研究鱼类的基因组结构、功能和进化规律的科学。
通过基因组测序和分析,科学家们可以揭示鱼类的遗传特征、物种多样性和进化历程。
这有助于我们更好地理解鱼类的生物学特性,为开发新型育种技术提供基础数据。
三、基因组育种技术基因组育种技术是一种利用基因组信息进行品种改良的新技术。
该技术通过分析个体的基因组信息,预测其表型特征,从而选择具有优良性状的个体进行繁殖,以实现品种的改良。
与传统的育种方法相比,基因组育种技术具有更高的选择准确性和育种周期短的优势。
四、基因组育种技术的应用生长性能:通过基因组育种技术,我们可以选择具有优良生长性能的个体进行繁殖,从而提高后代的生长速度和产量。
抗病性:通过分析鱼的基因组,我们可以预测其对抗特定疾病的抵抗力。
选择具有较强抗病性的个体进行繁殖,可以提高整个群体的抗病能力。
环境适应性:利用基因组育种技术,我们可以选择对不同环境条件具有较强适应性的个体进行繁殖,从而提高鱼类在不同水域环境中的生存能力。
品质改良:通过基因组育种技术,我们可以选择具有优良品质特征的个体进行繁殖,如肉质口感、营养价值等。
这有助于提高水产品的市场竞争力。
五、前景展望随着鱼类基因组学及基因组育种技术的不断发展,我们有望在未来实现更高水平的鱼类品种改良。
通过深入挖掘鱼类的遗传潜力,我们将能够培育出更具生长优势、抗病能力强和环境适应性好的新型鱼类品种。
这不仅有助于提高水产养殖业的生产效率和经济效益,还有利于保护生态环境和维护生物多样性。
六、结语鱼类基因组学及基因组育种技术是当前水产养殖领域的研究热点。
这些技术的不断发展将为我们提供更多有关鱼类生物学特性的深入了解,并推动水产养殖业的持续发展。
鱼类遗传育种的理论与实践研究

鱼类遗传育种的理论与实践研究鱼类育种是一门重要的学科,在农业和渔业中都有着重要的应用价值。
随着遗传学和分子生物学的发展,鱼类遗传育种研究也迎来了一个新的时期。
本文将从理论与实践两方面来探讨鱼类遗传育种的研究现状与前景。
一、理论研究1. 遗传多样性研究遗传多样性研究是鱼类遗传育种的基础。
通过对不同种群、不同品系间遗传变异的比较分析,可以筛选出优异的基因性状,为育种提供素材。
目前,对一些经济上重要的鱼类,如鲤鱼、鲢鱼等,进行了大量的遗传多样性研究。
不过由于鱼类种类繁多,现有研究仍然无法涵盖所有鱼类。
2. 基因组学研究基因组学研究是鱼类遗传育种领域中一个十分热门的话题。
通过对鱼类基因组的深入研究,可以揭示鱼类生长发育、抗病能力、耐逆性等重要性状的基因调控机制,为鱼类育种提供更为精确的基因选择工具。
现在,欧洲褐鳕和斑节鲆的基因组组装工作已经基本完成,而其他一些经济上重要的鱼类,如大马哈鱼、草鱼、青鱼等,也都有了初步的基因组序列资料。
3. 遗传改良理论研究鱼类遗传改良理论研究主要包括配套育种选择方法、定量遗传分析、遗传效应分析等方面。
这些理论研究对于鱼类种质改良和良种选育具有重要意义。
近年来,越来越多的遗传改良理论研究成果被应用在实践中,取得了不俗的成绩,如对多个品系鲤鱼进行了选代,产生了多个高生长速度的品系。
二、实践研究1. 选育优良鱼类品系选育优良鱼类品系是鱼类遗传育种的核心任务之一。
通过对经济上重要的鱼类进行长期、系统、细致的育种选择,可以培育出高生长速度、高产、高抗病性的高效鱼种,提高养殖效益。
目前,鲤鱼、鲢鱼、青鱼、鳙鱼、马鲅鱼等品种在优良品系选育方面研究较为深入,并取得了不错的效果。
2. 适应性育种研究适应性育种研究是应对气候变化等逆境环境的有效方式之一。
通过对鱼类的耐高温、耐酸碱度、耐盐碱性等逆境能力的研究,可以培育出具有高适应性的优良品系,提高养殖的稳定性和可持续性。
目前,一些适应性育种研究成果已成功应用于生产实践中,如南方红鲤鱼抗低氧适应性育种。
分子设计育种发展现状

分子设计育种发展现状
目前,分子设计育种是农业领域的一项新兴技术,它利用基因组学、生物信息学、计算机科学等多学科知识,通过预测、设计和优化植物或动物基因组以实现优良品质和高产量的育种目标。
与传统育种方法相比,分子设计育种具有更高的效率和精确性。
在分子设计育种的发展过程中,研究者们主要面临以下几个挑战。
首先,基因组测序技术的发展是分子设计育种的基础。
随着测序技术的不断提升,我们能够更准确地了解生物的基因组信息,并根据这些信息进行育种工作。
目前,新一代测序技术的应用已经成为分子设计育种的关键步骤。
其次,数据处理和分析的方法也是分子设计育种中的重要一环。
大规模的基因组数据需要通过生物信息学和计算机科学的方法进行分析,以提取有用的遗传信息。
同时,数据的质量和准确性对育种结果也有很大影响,因此需要不断改进和优化分析方法。
另外,分子设计育种还需要建立模型和算法来进行基因组设计。
通过模拟、预测和优化基因组组合,可以快速筛选并生成具有优良性状的个体。
这需要结合生物学原理和数学方法,建立相应的计算模型,以解决育种中的复杂问题。
此外,分子设计育种还面临着伦理、法律和社会等方面的挑战。
对于转基因植物或动物的研发和推广,有关食品安全、环境保护等方面的问题需要得到合理解决。
同时,公众对于分子设计育种的态度和接受程度也是影响其发展的重要因素。
总体而言,分子设计育种在基因组学、生物信息学及计算机科学等领域的不断进展,为实现高产量和优质品种的培育目标提供了有力支持。
随着技术的发展和挑战的解决,分子设计育种有望在未来更广泛地应用于农业及其他领域,促进粮食安全和农产品质量提升。
鲑鱼的基因组学和遗传学研究

鲑鱼的基因组学和遗传学研究鲑鱼是重要的经济鱼类,也是广受欢迎的食材之一。
在过去的几十年中,科学家们对鲑鱼进行了广泛的基因组学和遗传学研究,旨在揭示其独特的遗传特征和进一步改良鲑鱼的生长性能、抗病能力和适应能力。
本文将重点讨论鲑鱼基因组学和遗传学研究的重要进展以及对鲑鱼产业的影响。
鲑鱼基因组学的研究主要集中在两个方面:基因组测序和功能基因组学。
通过对鲑鱼基因组的整体测序,科学家们得以揭示鲑鱼基因组的组成和结构,包括基因数目、基因序列以及遗传变异的模式。
这项工作为全面了解鲑鱼的遗传信息奠定了基础,并为后续的功能基因组学研究提供了重要的数据支持。
功能基因组学则通过研究基因的功能和表达方式,进一步解析鲑鱼的遗传机制和生物学特征。
其中,转录组学和蛋白质组学是功能基因组学的两个重要方法。
转录组学研究的是基因的转录活动,通过对鲑鱼不同组织和生长阶段的基因表达进行分析,科学家们可以发现与特定生物学过程相关的基因,并进一步研究其调控网络和作用机制。
蛋白质组学则研究基因产物——蛋白质的表达和功能,有助于揭示鲑鱼基因与蛋白质之间的关系,以及它们在生理和生化过程中的作用。
基于这些基因组学的研究成果,鲑鱼遗传学工作得以深入开展。
鲑鱼遗传学的主要目标是解析鲑鱼的遗传特征,并利用遗传改良的方法提高鲑鱼的经济性状和环境适应性。
遗传改良方法包括选择育种、杂交育种、转基因等。
选择育种是通过对个体表型特征进行评估,选取具有优良性状的个体进行繁殖,逐步改良鲑鱼的生长速度、抗病能力等性状。
杂交育种则是利用不同品系或种间鲑鱼的杂交优势,获得更强大的遗传多样性,提高鲑鱼的适应性和抗病能力。
转基因技术则是将外源基因导入鲑鱼的基因组中,使其具备特定的性状,如抗病能力、生长性能等。
除了遗传改良,鲑鱼基因组学和遗传学的研究还为鲑鱼产业的可持续发展提供了许多新思路。
例如,通过基因组学的研究,科学家们发现了与鲑鱼生长发育、免疫抗病、环境适应等相关的基因,有助于培育耐高温、耐低氧等环境胁迫的鲑鱼品种。
水产动物遗传育种学

水产动物遗传育种学
水产动物遗传育种学是研究水产动物遗传变异和育种的学科,涉及到水产动物的遗传基因、染色体、突变、育种策略、品种改良等内容。
该领域的研究对于提高水产动物的生产力和品质,促进渔业可持续发展具有重要意义。
在水产动物遗传育种学研究中,主要涉及到以下方面:
1. 遗传基因研究:研究水产动物的遗传基因结构、功能和表达机制,探究遗传因素对水产动物生长、肉质、抗病性等的影响。
2. 染色体研究:研究水产动物的染色体数目、形态、结构和组型特征,探究染色体在水产动物遗传变异中的作用。
3. 育种研究:通过选择、杂交和栽培技术等手段,培育出具有良好生产性能、抗病性、品质好的水产动物新品种。
4. 品种改良:通过对现有水产动物品种的改良,提高其生长速度、产量、肉质和抗病性等方面的表现。
5. 生物技术应用:应用基因编辑、转基因等技术,改善水产动物的遗传性状,提高生产力和品质。
水产动物遗传育种学的研究对于提高水产动物的生产力和品质,促进渔业可持续发展具有重要意义。
几个重要农业基础研究方向的探讨

改善品种适应性和延长品种使用寿命的有效途径。 今后 农 作 物 育 种 基 础 研 究 的 重 点 方 向 是:
($)群体、个体和分子水平上品质、产量、抗性、适应 性、资源高效利用等相关性状的遗传变异规律和遗 传调控机制研究,重点是数量性状的数量遗传学和 分子遗传 学 基 础;(!)围 绕 国 家 粮 食 安 全 与 社 会 可 持续发展的重大需求,利用现代分子生物学研究策 略和手段,在分子、细胞、个体等不同水平上,开展与 作物基因资源发掘、优异育种材料创制和优良品种 培育等领 域 相 关 的 基 础 和 应 用 基 础 研 究;(()应 用 高 通 量 的 功 能 基 因 组 技 术( 如 基 因 芯 片、转 录 因 子 组、蛋白组研究技术等),开展重要农艺性状(产量、 抗逆性、抗病虫性、营养高效和品质等)的功能基因 组(新基 因 克 隆 与 功 能 分 析 )和 比 较 功 能 基 因 组 研 究,以作物种质资源为基础克隆重要农艺性状新基 因并进行功能、表达和调控分析,剖析作物重要农艺 性状的 形 成 机 制;())综 合 运 用 基 因 组 学、转 录 组 学、蛋白质组学和代谢组学信息,预测和认识生物体 各种性状的代谢、发育、分化和进化规律,为进行蛋 白质、核酸的分子设计或品种设计提供信息平台,研 究内容包括获取各种作物完整的基因组,建立主要 农作物重要农艺性状基因的数据库和信息系统,发 展基于功能基因信息的作物品种分子设计软件和方 法等; (#)杂种优势利用的基础理论研究,建立完善 杂种优势利用的育种理论和技术方法体系,实现育 种理论和技术研究的重大突破,加强两系杂种优势 利用的基础研究,研究光温敏不育的遗传学基础,构
此外,许多人畜共患的重大疫病,如血吸虫病、 狂犬病、乙型脑炎、链球菌病、流感等均与动物的带 毒、带菌、带虫有关,使这些病在动物和人之间相互 传播,对 人 类 身 体 健 康 构 成 严 重 威 胁。),,. 年 /01/的爆发 给 我 国 乃 至 全 球 经 济 造 成 巨 大 损 失; ),,2年初在亚洲禽流感严重爆发的情况下,我国先 后有!*个省(市、自治区)发生禽流感,给畜牧经济 和饲料工业带来了重大打击。联合国粮农组织总干 事雅克·迪乌夫在参加曼谷粮食安全问题地区会议 时表示,禽流感严重打击了亚洲国家的家禽养殖业, 不但对家禽养殖业造成了“灾难性”的打击,而且对 粮食安全和食品安全构成了威胁。因此加强重大畜 禽疫病的基础理论和防治技术的研究具有特殊重要 的意义。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
项目名称: 重要养殖鱼类功能基因组和分子设计育种的基础研究首席科学家: 桂建芳 中国科学院水生生物研究所 起止年限: 2010年1月-2014年8月依托部门: 中国科学院 湖北省科技厅一、研究内容基于拟解决的鱼类分子设计育种的策略和理论基础以及鱼类分子设计育种的可行性途径这两个关键科学问题,本项目拟集中在鱼类主要经济性状如生殖、生长、抗性的功能基因组和分子设计育种的基础方面,重点解析这些主要经济性状的基因调控网络,开拓关键技术,其主要研究内容包括:1)鱼类生殖基因调控网络及其分子设计育种的基础研究采用基因转移、morpholino介导的基因敲降等技术,配合整体原位杂交、细胞示踪和免疫荧光定位等方法,重点解析鱼类生殖质(germ plasm)的基因调控网络;揭示重要生殖调控基因在鱼类卵母细胞成熟和卵-胚转换中的功能作用及其调控机制;探讨鱼类配子发生过程中双亲特异性甲基化在生殖调控中的作用及其及其调控网络;通过大规模RFLP分析等呈现技术,筛选雌雄个体间性别特异的DNA片段标记,进而通过Genomic Walking克隆分离与性别决定和性别分化相关基因;揭示鱼类性别决定和性别分化的基因调控网络及其作用机理;筛选鉴定出可用于鱼类分子设计育种的性别特异表达基因和分子标记;开拓生殖调控基因和分子标记用于鱼类分子设计育种的可行性途径。
2)鱼类生长的基因调控网络和分子设计育种的基础研究采用morpholino介导的基因敲降、整体原位杂交、细胞示踪、免疫荧光定位以及组织碎片灌流和细胞孵育等在体和离体研究等技术,重点解析鱼类下丘脑/垂体控制生长的基因调控网络;揭示鱼类性成熟和生长的相互协调及其调控机理;探讨脑肠肽/生长激素/类胰岛素生长因子信号通路及其对鱼类生长的调控机制;揭示卵泡抑素等抑肌素相关基因在调控鱼类肌肉细胞增殖和鱼肉蛋白/脂肪平衡的作用机理;研制可控不育性转卵泡抑素等基因的转基因鱼品系;阐明鱼类个体大小调控基因的系统发育和进化规律;开拓生长基因调控网络和分子标记尤其是主控基因用于鱼类分子设计育种的可行性途径。
3)鱼类抗性的基因调控网络和分子设计育种的基础研究利用我们自主建立的分离鱼类抗病毒基因的细胞模型,重点解析鱼类干扰素系统和抗菌肽基因的调控网络和抗病作用机理;揭示重要鱼类病原与宿主免疫系统的相互作用及其作用机制;探讨在鱼类抗病免疫反应中主效基因调控网络、功能和抗病机理;研制具有抗病性状的雌核发育系和近交家系;鉴别出与鱼类抗病性状密切相关的分子标记和等位基因及其抗病QTL定位;建立鱼类抗病分子设计育种的理论和技术方法,开拓其可行性途径。
采用大规模测序和基因芯片分析等手段,通过比较转录组学和基因功能分析等研究手段,解析鱼类抗寒基因调控网络及其作用机理,揭示低温胁迫下不耐寒鱼类基因表达谱与耐寒鱼类基因表达谱的差异及其关键基因的功能;创制鱼类抗寒机理研究模型,开拓鱼类抗寒基因用于分子设计育种的可行性途径。
4)鱼类分子设计育种的关键技术研究通过建立鱼类数量性状基因调控网络信息整合技术和鱼类经济性状主效基因的图谱定位,揭示鱼类经济性状主效基因同线性和性状关联性关系,阐明鱼类经济性状主效基因转录组以及它们在群体和家系中的功能,确定鱼类经济性状主效基因标记和基因不同组合的遗传效应,评估其育种效率,由此创建可用于鱼类分子设计育种的数量性状多基因聚合育种的技术路线。
通过创制基于蛋白分子设计、BAC克隆、转座子系统、Cre/lox基因靶位操作的稳定、高效的鱼类转基因技术,优化和完善基于蛋白质锌指结构的鱼类基因快速敲除系统,创建基于破坏原生殖细胞发生或维持的具有遗传安全的鱼类转基因技术体系等建立可用于鱼类分子设计育种的基因操作技术路线,并揭示性腺特异小RNA在基因表达调控中的作用,阐明piRNA与互作蛋白的调控途径,研制出持续激活的生长激素受体基因的转基因鱼,揭示持续转激活生长激素受体基因与转生长激素基因的转基因鱼的效应及其信号通路的调控差异。
二、预期目标本项目的总体目标:针对水产养殖业发展现状和可持续发展需求,通过开展重要养殖鱼类生殖、生长和抗性等主要经济性状的功能基因组研究,解析鱼类生殖的基因调控网络、鱼类生长的基因调控网络、鱼类抗病和抗寒的基因调控网络,筛选鉴定进行分子设计育种的主控或关键基因和分子标记,建立鱼类分子设计育种的关键技术及其技术体系,在理论上阐明鱼类生殖、生长、抗病和抗寒等主要经济性状和重要生命现象的基因调控网络及其作用机理,提出鱼类良种分子设计的策略;在技术方法上,建立鱼类分子设计育种的多基因聚合和基因操作技术,创制优质育种材料,创建鱼类分子设计育种的可行性途径。
通过这些研究,使我国在鱼类功能基因组研究中居国际领先水平,为培育高产优质养殖新品种奠定理论和技术基础,为我国水产养殖业的可持续发展和渔业生物技术的创新做出贡献。
五年预期目标:1.解析鱼类生殖、生长、抗病和抗寒的基因调控网络;2.筛选鉴定具有自主知识产权、可用于鱼类分子设计育种的功能基因和分子标记30-50个;3.揭示鱼类性别决定与分化、卵母细胞成熟和卵-胚转换的分子机制;4.阐明鱼类生长激素分泌的信号通路;5.阐明鱼类干扰素系统关键基因和重要抗菌基因的抗病功能及其作用机理;阐明鱼类抗寒关键信号通路及其作用机理;6.建立鱼类数量性状多基因聚合育种技术;7.创建基于蛋白分子设计和遗传安全的稳定高效的基因操作技术;8.建立主要经济性状功能基因组研究和分子设计育种中间的有机联系,创制2-3个在生殖、生长 或抗性等目标经济性状上表现优质的育种材料;9.凝聚和培养一批有国际影响的中青年学术带头人和学术骨干;发表一批高质量的学术论文,其中在有重要国际影响的期刊(相关领域引用排名前15%的期刊或影响因子大于5的期刊)发表论文50篇以上。
三、研究方案(一)、学术思路:针对水产养殖业发展现状和可持续发展需求,以鲫和鲤等为主要研究对象,启动我国重要养殖鱼类功能基因组和分子设计育种的基础研究,通过研究鱼类生殖、生长和抗性的基因调控网络及其作用机理,筛选鉴定进行分子设计育种的关键基因和分子标记,建立鱼类分子设计育种的关键技术及其技术体系,解决鱼类分子设计育种的策略和理论基础以及鱼类分子设计育种的可行性途径这两个关键科学问题,为鱼类分子设计育种的实践和应用以及为我国水产养殖业的可持续健康发展和渔业生物技术的创新做出贡献。
(二)、技术途径:1. 采用solexa深度测序等大规模测序和基因芯片分析等手段,通过比较转录组学分析,了解鱼类生殖和生长不同阶段、以及抗病和不抗病、耐寒和不耐寒的基因表达谱差异,经生物信息学比较和分子特征分析,由此筛选调控鱼类生殖、生长、抗性的主要基因和关键基因;2. 建立重要鱼类基因组BAC库,研究基因的结构,克隆分离相应基因的启动子;将其启动子与GFP报告基因重组融合,利用基因转移等技术,采用流式细胞仪等筛选分析技术,进一步研究生物学功能;3. 建立鱼类BAC转基因技术,用杂交方法或PCR方法筛选重要功能基因的BAC克隆,在基因编码框的适当位置插入绿色荧光蛋白(EGFP)基因;利用以上构件制备转基因鱼,通过EGFP分析这些基因的时空表达情况及其生理学效应;4. 通过基因连锁分析和原位杂交等技术,研究目的基因在染色体上的定位,揭示基因相互作用的基因组组织基础;5. 用RT-PCR、原位杂交和Western blot方法在mRNA及蛋白水平上检测基因表达的组织特异性和时序性;用免疫荧光定位确定其基因的蛋白产物的组织、细胞和亚细胞定位;6. 采用染色质免疫沉淀法(Chromatin immunoprecitation,ChIP)、凝胶迁移或电泳迁移率检测(Electrophoretic Mobility Shift Assay,EMSA)等技术研究DNA 与蛋白的相互作用;7. 采用免疫共沉淀(Coimmunoprecipitation, Co-IP)、GST-pull down、荧光共振能量转移(Fluorescence resonance energy transfer,FRET)、双分子荧光互补(bimolecular fluorescence complementation,BiFC)等技术研究蛋白相互作用;对一些重要蛋白进行质谱分析、并对其进行泛素化、乙酰化和磷酸化等修饰分析;8. 鉴定两性生殖鲫鱼卵子中双亲特异性甲基化、发育不等性基因,分析单性生殖鲫鱼三倍体卵子中相关同源基因启动子CpG岛甲基化差异和基因表达调节差异,确定发生差异甲基化的目标基因;比较分析两性生殖与单性生殖鲫鱼卵子基因组中差异甲基化功能基因启动子,鉴定差异甲基化顺式调控序列。
9. 采用正常的和人工突变后的基因转移试验,辅以其它分子生物学方法如目的基因表达产物体内、外诱导处理等来分析目的基因的生理功能;10. 用显微注射方法研究基因功能。
将含有这些基因的真核表达载体,或在大肠杆菌中表达出的蛋白,注射到胚胎中或转染到细胞中,观察生物学效应;11. 将目的基因的反义RNA或其蛋白的抗体注射到胚胎中,观察生物学效应;12. 对生殖调控基因而言,设立人工性反转实验,分别提取不同性反转阶段实验组和对照组性腺的mRNA和蛋白,采用上述方法研究基因表达的时序性;13. 对生长和生殖功能基因而言,合成重要调控生长生殖基因的结构类似物或表达具有生物活性重组蛋白,取鱼类的下丘脑、垂体、肝脏和性腺,通过碎片灌流和细胞孵育等离体研究方法,或直接注射结构类似物或重组蛋白,采用放射免疫测定等方法研究其活性,测定各种靶组织、血清中调控生殖生长激素的含量,分析这些生长调控因子和生殖调控因子的相互作用及其信号传递机制,分析它们对鱼类生殖和生长的影响;14. 通过体外合成小分子RNA,建立鱼类系统中适用于实验室日常工作的、简洁、高效的siRNA技术平台;通过克隆获得的鱼类来源的启动子构建针对草鱼出血病病毒的siRNA载体,建立抗病转siRNA基因鱼模型;通过Cre-loxp介导的系统进行鱼类胚胎中的条件基因打靶研究;建立鱼类胚胎干细胞和生殖干细胞系,建立鱼类培养细胞体外基因操作和个体重建技术途径;15. 通过morpholino介导的基因敲降技术,配合整体原位杂交、细胞示踪等方法,研究目的基因的生理学功能及其基因调控网络、信号通路和作用机理等;16. 采用已经建立的细胞模型,研究病毒与宿主细胞的相互作用,在此基础上,通过转染目的基因和siRNA干扰等技术手段,研究目的基因的生理学功能以及基因的调控网络;17. 通过AFLP和微卫星等遗传标记的大规模筛选,鉴定出与鱼类性别等等主要经济性状相关的分子遗传标记,进而通过Genomic walking克隆鉴定相关功能基因;通过人工选育和家系分析,利用遗传标记进行基因型分析的优势,研究特定标记与某些经济性状(例如生长、性别、抗病或抗寒)的连锁关系,进行分子标记辅助育种;开展鱼类生长、饲料转化率等经济性状的QTL定位分析和eQTL定位分析;18. 改造基于蛋白质锌指结构的基因敲除载体系统,优化基因敲除载体,引入更多的克隆位点,以简化特异基因敲除载体的构建;利用Zinc Finger Tools 网上软件包(),选取代表性基因进行锌指结构域的索寻,选择连续20或30个氨基酸的合适结构域,依据针对DNA三联体的经验锌指氨基酸序列,设计靶标多肽和碱基序列;将靶标多肽的DNA片段克隆到经改进后的基因敲除载体上;体外合成mRNA,显微注射;观察表型,并利用PCR检测显微注射个体的基因型,获得杂合体;将杂合体个体扩大培养,并进行相互交配,以获得基因的两个拷贝均缺失的纯合体;系统分析基因缺失纯合体的表型。