操纵子1
每周例题-操纵子--生物奥赛

1.关于操纵基因的叙述,下列那项是正确的:A.与阻遏蛋白结合的部位B.与RNA聚合酶结合的部位C.属于结构基因的一部分D.具有转录活性E.促进结构基因转录2、为什么葡萄糖可参与乳糖操纵子的代谢阻遏?A.与乳糖操纵子的调控毫无关联B.葡萄糖的降解产物能降低细胞内cAMP的含量C.因为葡萄糖也是β-半乳糖苷酶的底物D.葡萄糖的存在增加了细胞内乳糖阻抑物的含量3、色氨酸操纵子的终产物——色氨酸如何参与操纵子的调控?A.结合到阻抑物上,阻断其与DNA 的结合,从而使转录得以进行B.结合到阻抑物上,使阻抑物与DNA 结合,从而使转录得以进行C.色氨酸直接与DNA 结合,抑制操纵子转录D.结合到阻抑物上,形成复合物与DNA 结合,阻止转录的进行这周为大家分享的是一些有关于操纵子的题目,因为近年来,操纵子确实是一个常考点,况且由于今年分析类题目的比重上升,因此操纵子这类需要理解的题目考核的可能性再一次上升。
下面进入今天的分析。
首先在讲操纵子之前,先了解操纵子的基本结构。
操纵子由启动子(P)、操纵基因(O)、以及结构基因组成,一定要注意,虽然调节基因(I)对操纵子起到调节作用,但它不属于操纵子它存在操纵子的上游。
调节基因表达阻遏蛋白,和各类底物结合,激活或抑制阻遏蛋白,对操纵子的表达起作用。
启动子是基因开始表达的起始位点。
操纵基因是阻遏蛋白的结合部位。
而结构基因就是能够表达出各种有生理功能的多肽的基因片段。
下面介绍两种常考的操纵子,分别是乳糖操纵子和色氨酸操纵子,希望大家了解之后,今后遇到相关题目能够做对。
乳糖操纵子当培养基没有乳糖时,调节基因表达出的阻遏蛋白和操纵基因结合,阻止了结构基因(一些转运和分解乳糖的酶)的表达。
当在培养基中只有乳糖时,由于乳糖的代谢产物异乳糖是lac操纵子的诱导物,它可以结合在阻遏蛋白的变构位点上,使构象发生改变,破坏了阻遏蛋白与操纵基因的亲和力,不能与操纵基因结合,于是RNA聚合酶结合于启动子,并顺利地通过操纵基因,进行结构基因的转录,产生大量分解乳糖的酶,这就是当大肠杆菌的培养基中只有乳糖时利用乳糖的原因。
Lecture1 乳糖操纵子

Ph.D.Quan Chen,2002‐2007, Ph.D. in Biochemistry and Molecular Biology, USTC,,y,,,2007‐2009, Research Fellow, Mayo Clinic, Scottsdale, AZ, USA2009‐2011, Research Associate, University of Southern California, Los Angeles, CA, USA 2011‐present, Associate Professor, USTCMolecular Biology第一章前言I 第二章基因和基因组第三章DNA复制第四章转录Undergraduate 第五章翻译第六章基因表达的调控第七章突变与修复第八章癌症的分子生物学GraduateII 第九章转座子第十章基因操作和基因组分析第十一章细胞核的分子生物学第十二章基因表达调控的分子机制III 第十三章疾病产生的分子基础第十四章基因诊断和基因治疗Molecular Biology II ¾Regulation of Gene Expression‐In Prokaryotes‐In Eukaryotes¾Mutation and RepairG ti f C¾Genetics of Cancer¾Transposable Elements¾Gene Manipulation & Genomic AnalysisReferences《Essential i Genetics》, Peter J Russell The Benjamin/Cummings Publishing Company, Inc 《Genetics》, 5th ed.,Peter J Russell The Publishing Company, ed Benjamin/Cummings Company Inc Genes IX, Benjamin Lewin Jan. 2008 ISBN: 9787030182982.Grading g30% ‐Attendance, homework and quizzes70% ‐Final examFi lWhy is Regulation necessary• the level of Not all genes are expressed continuously:gene expression differ according to‐‐cell types, stages of cell cycle•Organisms live in changing environments•Regulation allows organisms to grow and reproduce optimally in different environmentsti ll i diff t i tAre all genes’ activity regulated?•The housekeeping genes (constitutive genes)Genes that are essential for normal cell functionf fand are constitutionally expressedg g•The regulated genesTheir activity is controlled in response to the needs of a cell ororganism•Distinction between regulated and constitutivegenes is somewhat arbitraryRegulation of Gene ExpressionThe most direct control pointRegulate by changing sets of genes’ expression level in response to environment alterationi l iEfficiency and economicsRegulation of Gene Expression in Prokaryotes ¾The lac Operon of E.coli¾The trp Operon of E.coli¾Regulation of gene expression in Phage Lambdal f h bdThe Logic of Prokaryotic Gene RegulationEfficiency and economicsMetabolic enzymes are required to break down different carbon sources to yield energy.Carbon sources: lactose, glucose, galactose, xylose etc.Selective repression and activation mechanisms must fulfill two criteria:They must have the ability to reversibly turn on or switch off transcription of each specific gene or group of genes.They must be responsive to environmental conditions in which transcription of the relevant gene(s) should be activated or repressed.Prokaryotic transcriptionPromoter–recruit RNA PolymeraseOperator sites–bind activators/ repressors10¾‐Element¾‐35 Element¾Extended ‐10 Element¾UP elementKey ConceptsTwo types of DNA-p rotein interactions are required for regulated transcription. determines where transcription begins-Promoter-RNA Polymeraseyregulates whether or not promoter-driven transcription occursOperators-Operators-Activators/ repressorsgMessageGenes must contain two kinds of binding sites to permit regulated transcription. First, binding sites for RNA polymerase must be present. Second, binding sites for activator or proteins must be present in the vicinity of the promoter.repressorAllosteric effectorseffectors toggle switches to control the ability or Allosteric work like of activator repressor proteins to bind to their DNA target sites.Operons‐Fine Control of Prokaryotic TranscriptionThe genes, the operator, and the promoter constitute an operonOperon‐‐Genes grouped together; transcribed into single mRNA molecule containing coding sequences for more than one gene (polycistronic mRNA). Grouped together with one promoter.Structural gene–lacZ, Y, ARegulatory gene ‐lacIOperon. A cluster genes, their expression are coordinatelyof genesregulated by operator/regulator protein interactions, plus the operator and promoterregion the¾l lac operon¾trp operoncontrol–repressorNegative controlPositive control–activatorNegative and positive regulation are not mutually exclusiveLactose metabolism and the operonWhat led to the study of operon regulationNoDiauxiediauxieGlucose GalactoseMannose ArabinoseFructose XyloseMannitol RhamnoseSorbitolS bi lDulcitolMaltoseLactose•when cultures of E.coli were grown in mixtures of two different sugars. Depending on the particular sugarsinvolved, the resulting growth curve showed eitherinvolvedcontinuous or biphasic growthThey wanted to know•How bacteria know that lactose is present in the growth medium?•What role the inducer plays in enzyme synthesis?•How do the cells respond so rapidly to the presence of a new sugar?Bacterium’s adaptation to utilizing different nutrients •Glucose is the main source of carbon•Enzymes required for glucose metabolism are coded byconstitutive genesi i•In the absence of Glucose and presence of other carbonsources such as lactose,the genesfor utilizing these substance are ‘turned on”.•The related induciblegenes are:(regulated genes)•The alternate substance : inducer•the effectors that turn repressoroff these genes:The general structure of a inducible geneofMetabolism lactosesimplified lac operon model ‐galactosidase: cleave the lactose moleculeb l id l h l l l Permease: transport lactose into the cellTransacetylase: not required for lactose metabolismβ‐galactosidase function in two waysCarbon sourceInducerdRegulation of the lac operonPolycistronic mRNA: several genes are transcribed into one mRNA.To ensure: gene products required for the same event are produced equally.T d t i d f th t d d lllactose must be present in the environmentglucose cannot be present in the cell's environmentLac operon controlling regionNegative regulation of the lac operon•lac I gene encodes lac repressor•The repressor is an allosteric protein•Inducer: allolactose, IPTGand a low basal level•Repressor is leaky of the lac operon products is always present.•Only about 10 tetramers of repressor are present per cell.The repressor protein•tetramer•an allosteric protein with two domains:a domainDNA bindingan inducer binding domainInducer binding Æstructural change at DNA binding domain Ælose its DNA binding ability Ætranscription repression relieved.The lac operon repression model In absence of lactheIn the presence of lactoseinducer ‐allolactoseE i l id f h Experimental evidence for the regulation of lac genesMutation in the Protein‐Coding Genes‐‐‐‐polar effect on a polygenic mRNA•LacZ, LacY and Lac A mutation to map theirlocation•Missense mutation•Nonsense mutationIn sum, nonsense mutations in the cluster of three genes involved in lactose utilization have different effects, depending on where they are located within the cluster: The nonsense mutations are said to exhibit polar effects, and the phenomenon is called polarity. (nonsense mutations that show polar effects therefore are called polar mutation)Mutation affecting the Regulation of Gene ExpressionConstitutive Mutation: Mutation that cause its downstream genes constitutively C i i M i M i h i d i i lexpressed with or without inducerTwo groups of constitutive mutations:Operator Mutation‐‐‐‐‐‐mapped to a small region near lac Z(lac O)LacI Gene Regulatory Mutation‐‐‐‐‐‐mapped to a DNA region a small distance away (lac I)‐‐repressor geneThe Operator Mutations•Partial diploid: F’lacO+ lacZ‐lacY+‐lacO c lacZ+ lacYg•F’strains in which a few chromosomal genes on an extrachromosomal genetic element called F factor are introduced into a bacterial cell•Test for gene products both in the presence and the absence of inducer allolactoseb f i d ll lβ•in the absence of inducer: ‐galactosidase•in the presence of inducer: β‐gal and permeaseCis‐dominance•Partial diploid: F’lacO+ lacZ‐lacY+‐lacO c lacZ+ lacYp g q y •The phenomenon that a gene or DNA sequence only controlling structure genes on the same, contiguous of DNA‐piece cis dominance•generally speaking, a cis‐dominant element has no diffusible gene product (protein)diff ibl d(i)•lacO+/lacO c on different chromosomes do not interfere. And has no position effectlacI gene mutations•Partial diploid: F’lacI+lacO+lacZ‐lacY+‐++‐lacI lacO lacZ lacY•Test for structural gene expression in the presence and the absence of inducerIn no βgal and permease•the absence of inducer:‐•in the presence of inducer: both proteins were made •The lacI+ gene can overcome lacI‐defect. Even when they are located on different chromosomesTrans‐dominance•Partial diploid: F’lacI+lacO+lacZ‐lacY+lacI‐lacO+lacZ+lacY‐•A gene or DNA sequence that can control genes on different, noncontiguous piece of DNA,g p•A trans‐dominant element must produce a diffusible geneproduct(protein)•In one system, a lacI+ gene can overcome any lacI‐gene defect.defectmutations ‐‐‐‐‐‐Cis‐dominantlac promoter()g •Promoter mutations Plac‐): affect all three structural genes (none or few), even with inducer present•Plac‐only control genes that are on the same DNA strand‐‐‐Cis‐dominantEffect of MutationsOperator mutationsLacO c mutation: operator DNA sequence mutatesp qcould not be recognized by the repressorits downstream genes constitutively expressed(with or inducer)withoutPartial diploid in the absence of inducerNoexpressionConstitutiveexpressionPartial diploid in the presence of inducerInducibleexpressionconstitutiveexpressionHaploid in presence or absence of inducerconstitutiveexpressionu a ep esso cou d o b d o ope a oMutant repressor could not bind to operatorPartial diploid in the absence of inducerNoexpressionPartial diploid in the presence of inducerInducibleexpressionInducibleexpression。
操纵子

操纵子(operon):指启动基因、操纵基因和一系列紧密连锁的结构基因的总称。
转录的功能单位。
很多功能上相关的基因前后相连成串,由一个共同的控制区进行转录的控制,包括结构基因以及调节基因的整个DNA序列。
主要见于原核生物的转录调控,如乳糖操纵子、阿拉伯糖操纵子、组氨酸操纵子、色氨酸操纵子等1961 年 ,法国巴斯德研究所的 Monord 和Jacob 提出了乳糖操纵子概念 ,后来人们在大肠杆菌中又陆续发现了色氨酸操纵子、组氨酸操纵子、半乳糖操纵子、阿拉伯糖操纵子等多种操纵子 ,从而不断的充实和完善了被誉为生命第三原理的基因调控理论 ,在这个理论中提出的操纵子概念也被人们普遍接受和证实。
操纵子学说是关于原核基因结构及其表达调控的学说,由法国巴斯德研究所著名科学家的Monod和Jacob在1961年首先提出[1]。
他们以对乳糖操纵子的研究,通过大量的试验及分析建立了现在已经被人们广泛接受的乳糖操纵子控制模型[2]。
后来人们在大肠杆菌中又陆续发现了色氨酸操纵子、组氨酸操纵子、半乳糖操纵子、阿伯糖操纵子等多种操纵子,从而不断的充实和完善了被誉为生命第三原理的基因调控理论,在这个理论中提出的操纵子概念也被人们普遍接受和证实。
操纵子学说:1961年,法国科学家莫诺(J·L·Monod,1910-1976)与雅可布(F·Jacob)发表“蛋白质合成中的遗传调节机制”一文,提出操纵子学说,开创了基因调控的研究。
四年后的1965年,莫诺与雅可布即荣获诺贝尔生理学与医学奖。
莫诺与雅可布最初发现的是大肠杆菌的乳糖操纵子。
这是一个十分巧妙的自动控制系统,这个自动控制系统负责调控大肠杆菌的乳糖代谢。
乳糖可作为培养大肠杆菌的能源。
大肠杆菌能产生一种酶(叫做“半乳糖苷酶”),能够催化乳糖分解为半乳糖和葡萄糖,以便作进一步的代谢利用。
编码半乳糖苷酶的基因(简称z)是一个结构基因(structural gene)。
《阿拉伯糖操纵子》课件

阿拉伯糖操纵子与其他基因表达调控元件的相互作用
转录因子
研究阿拉伯糖操纵子如何受到转录因 子的调控,以及转录因子如何与其他 调控元件相互作用,共同调节基因的 表达。
染色质重塑
探讨阿拉伯糖操纵子如何通过染色质 重塑来影响基因的表达,以及染色质 重塑与其他调控机制之间的相互作用 。
05
阿拉伯糖操纵子的未来展望
当环境中存在乳糖时,乳糖会与阻遏蛋白结合,使其失去活性,解除对结构基因的 抑制,诱导相关酶的合成。
此外,阿拉伯糖操纵子还受到葡萄糖的抑制,当细胞内葡萄糖浓度过高时,会通过 未知机制抑制环腺苷受体蛋白的活性,从而抑制结构基因的表达。
02
阿拉伯糖操纵子的调控机制
阿拉伯糖的诱导作用
阿拉伯糖的诱导作用是指当阿 拉伯糖存在时,能够诱导阿拉 伯糖操纵子表达的现象。
通过调节阿拉伯糖操纵子的表达,可以控制微生物的代谢产 物,从而生产出具有特定活性或功能的药物。例如,利用阿 拉伯糖操纵子调控微生物的次级代谢产物,可以生产抗生素 、抗癌药物等。
阿拉伯糖操纵子在生物燃料生产中的应用
生物燃料是指利用生物质资源生产的燃料,如乙醇、生物 柴油等。阿拉伯糖操纵子在生物燃料生产中也有着重要的 应用。
阿拉伯糖操纵子在解决全球性挑战中的潜在作用
粮食安全问题
通过研究和利用阿拉伯糖操纵子 ,可以开发出抗逆、抗病、高产 的转基因作物,提高粮食产量和 质量,为解决全球粮食安全问题
提供有力支持。
气候变化问题
阿拉伯糖操纵子的研究可以为生 物固碳和减排提供新的思路和方 法,有助于减缓气候变化的影响
。
人类健康问题
转录活性。
在没有阿拉伯糖的情况下,阻遏 蛋白与操纵序列结合,导致操纵
子基因的表达受到抑制。
6-1阿拉伯糖操纵子

4. However genetically both AraI and AraO2 needed to inhibit BAD expression - proposed that Ara C protein binds simultaneously to AraI and AraO2.
When arabinose present, BAD expression is enhanced but C-protein synthesis still repressed. (absence of glucose, hi cAMP) a. Under this condition still see footprinting at same three sites of DNA. Found under this condition, CAP site is also bound with CAP protein and cAMP (so Cprotein can act either as an repressor or an activator depending on presence of arabinose.) ) b. Analysis of the DNA structure indicates C-protein forms a loop in the DNA when the CAP site is not filled.
的碳源,可以诱导ara ara操纵子 阿拉伯糖作为E.Coli的碳源,可以诱导ara操纵子 的转录。当细胞中葡萄糖水平高(导致cAMP处于低浓度) 的转录。当细胞中葡萄糖水平高(导致cAMP处于低浓度) cAMP处于低浓度 和阿拉伯糖水平低时,AraC阻遏araB,araA,araD的转录。 和阿拉伯糖水平低时,AraC阻遏araB,araA,araD的转录。 阻遏araB 的转录
操纵子

1?乳糖操纵子的结构大肠杆菌的乳糖操纵子含Z、Y及A三个结构基因,分别编码β-半乳糖苷酶、透酶、乙酰基转移酶,此外还有一个操纵序列O、一个启动序列P及一个调节基因Ⅰ。
Ⅰ基因编码一种阻遏蛋白,后者与O序列结合,使操纵子受阻遏而处于转录失活状态。
在启动序列P上游还有一个分解(代谢)物基因激活蛋白CAP 结合位点,由P序列、O序列和CAP结合位点共同构成LAC操纵子的调控区,三个酶的编码基因即由同一调控区调节,实现基因产物的协调表达。
2?阻遏蛋白的负性调节在没有乳糖存在时,乳糖操纵子处于阻遏状态。
此时,Ⅰ基因列在P启动序列操纵下表达的乳糖阻遏蛋白与O序列结合,故阻断转录启动。
阻遏蛋白的阻遏作用并非绝对,偶有阻遏蛋白与O序列解聚。
因此,每个细胞中可能会有寥寥数分子β半乳糖苷酶、透酶生成。
当有乳糖存在时,乳糖操纵子即可被诱导。
真正的诱导剂并非乳糖本身。
乳糖经透酶催化、转运进入细胞,再经原先存在于细胞中的少数β -半乳糖苷酶催化,转变为别乳糖。
后者作为一种诱导剂分子结合阻遏蛋白,使蛋白构型变化,导致阻遏蛋白与O序列解离、发生转录,使β-半乳糖苷酶分子增加1000倍。
3?CAP的正性调节分解代谢物基因激活蛋白CAP是同二聚体,在其分子内有DNA结合区及cAMP结合位点。
当没有葡萄糖及cAMP浓度较高时,cAMP与CAP结合,这时CAP结合在乳糖启动序列附近的CAP位点,可刺激RNA转录活性,使之提高50倍;当葡萄糖存在时,cAMP浓度降低,cAMP与CAP结合受阻,因此乳糖操纵子表达下降。
由此可见,对乳糖操纵子来说CAP是正性调节因素,乳糖阻遏蛋白是负性调节因素。
两种调节机制根据存在的碳源性质及水平协调调节乳糖操纵子的表达。
4?对调节机制的解释大肠杆菌根据碳源性质选择代谢方式。
倘若有葡萄糖存在时,细菌优先选择葡萄糖供应能量。
葡萄糖通过降低cAMP浓度,阻碍cAMP与CAP 结合而抑制乳糖操纵子转录,使细菌只能利用葡萄糖。
典型乳糖操纵子的诱导原理

操纵子(operon):很多功能相关的结构基因串联排列在染色体
上,由一个共同的控制区来操纵这些基因的表达,包含这些结构基 因和控制区的整个核苷酸序列就称为操纵子。
乳糖操纵子的结构和功能
操纵子的组成: ----结构基因(structural gene, SG) :操纵元中被调控的编码蛋白质的 基因 ----启动子(promoter,P):是指能被RNA聚合酶识别、结合并启动基 因转录的一段DNA序列。 ----操纵基因(operator,O):是指能被调控蛋白特异性结合的一段 DNA序列。 阻遏物基因(inhibitor,I),产生阻遏物(repressor)。
乳糖操纵子的结构和功能
结构基因 • Z编码β-半乳糖苷酶:将乳糖水解成葡萄糖和半乳糖。 • Y编码β-半乳糖苷透过酶:使外界的β-半乳糖苷(如乳糖) 能透过大肠杆菌细胞壁和原生质膜进入细胞内。 • A编码β-半乳糖苷乙酰基转移酶:乙酰辅酶A上的乙酰基转 到β-半乳糖苷上,形成乙酰半乳糖。
乳糖操纵子的结构和功能
乳糖操纵子的调控机理
乳糖操纵子诱导物
是别 诱乳 导糖 物
(allolactose)
为什么选用IPTG作诱导物?
能诱导酶的合成,但又不被分解的分子,称为安慰 诱导物(gratuitous inducer)。 由于乳糖虽可诱导酶的合成,但又随之分解,产 生很多复杂的动力学问题,因此人们常用安慰诱 导物来进行各种实验。 X-gal(5-溴-4-录-3-吲哚-β-半乳糖苷)也是一种 人工化学合成的半乳糖苷,可被β-半乳糖苷酶 水解产生兰色化合物,因此可以用作β -半乳糖 苷酶活性的指示剂。IPTG和X-gal都被广泛应用 在分子生物学和基因工程的工作中。
典型乳糖操纵子的诱导原理

典型乳糖操纵子的诱导原理乳糖操纵子是一种在分子生物学和遗传工程领域广泛应用的工具。
它是由乳糖操作子(lac operator)和乳糖调节因子(lac repressor)组成的。
乳糖操作子是原核生物中的一段DNA序列,它位于乳糖操纵子基因(lac operon)的上游。
乳糖操作子可以结合乳糖调节因子,从而调控乳糖操纵子基因的转录。
乳糖调节因子是一种DNA结合蛋白,它分为两个亚单位。
乳糖调节因子通过与乳糖操作子的结合,可以起到两种不同的作用。
首先,当乳糖调节因子与乳糖操作子结合时,它会阻止RNA聚合酶结合到乳糖操纵子基因的启动子区域。
这意味着乳糖操纵子基因的转录被抑制,表达水平较低。
其次,当乳糖存在于细胞内时,它可以与乳糖调节因子结合并改变其构象。
这种构象改变会导致乳糖调节因子与乳糖操作子的结合断裂,从而释放乳糖操纵子基因的启动子区域。
这使得RNA聚合酶能够结合到启动子区域上进行转录,从而使乳糖操纵子基因得以高效转录,表达水平增加。
总结起来,乳糖操纵子的诱导原理可以归纳为两个方面。
一方面,乳糖调节因子与乳糖操作子的结合可以抑制乳糖操纵子基因的转录,从而控制其表达水平。
另一方面,乳糖存在时,它与乳糖调节因子结合并改变其构象,使乳糖调节因子与乳糖操作子的结合断裂,从而释放启动子区域,促进乳糖操纵子基因的转录。
乳糖操纵子的诱导原理在实验室中被广泛应用于表达外源基因。
通过将目标基因与乳糖操纵子基因合并,将其导入细菌或其他原核生物中,可以实现对外源基因的可控表达。
当乳糖存在时,外源基因能够高效转录,并表达出目标蛋白。
而当乳糖缺失时,乳糖调节因子与乳糖操作子的结合会导致转录抑制,从而停止对外源基因的表达。
乳糖操纵子的诱导原理不仅在实验室中被广泛应用,也在工业生产中发挥重要作用。
例如,在生物制药中,乳糖操纵子可被用来调控特定基因的表达水平,从而生产大量的目标蛋白。
此外,乳糖操纵子也可以用于研究基因调控网络和药物研发等领域。
操纵子模型

学 专
生: 业:
指导老师:
日
期:2
主要内容
一、操纵子系统的组成 二、结构基因的转录 三、乳糖操纵子的诱导表达模型
一、操纵子系统的组成
二、结构基因的转录
图(a) 表示由一个启动子、一个结构基因和终止子构成的一个操纵子。 开始位点和终止点影响转录
二、结构基因的转录
图(b) Sigma(σ)因子(蛋白质)与RNA聚合酶结合并刺激聚合酶与启动 子位点结构。没有σ因子,就不能有效地结合。
三、乳糖操纵子的诱导表达模型
三、乳糖操纵子的诱导表达模型
cAMP受体蛋白 结合位点 RNA聚合 酶结合位 点 阻抑物结 合位点 乳糖通透酶 β半乳糖苷 酶 乳糖转乙 酰酶
图3-1 乳糖操纵子
三、乳糖操纵子的诱导表达模型
cAMP 阻抑物蛋白
cAMP
cAMP 受体蛋 白
图3-3 E-Coli在缺葡萄糖,含有乳糖,cAMP充足,转录
二、结构基因的转录
图(c) RNA聚合酶开始转录后不久,sigma因子就与该酶分离。
二、结构基因的转录
图(d) 当RNA聚合酶到达终止位点,转录停止;Rho因子(蛋白质)与 RNA聚合酶结合并促进该酶与DNA分离和RNA的释放
二、结构基因的转录Fra bibliotek图(e) RNA聚合酶、Rho因子和从DNA分离出来的新 生的RNA
三、乳糖操纵子的诱导表达模型
阻抑物蛋白
cAMP cAMP 受体蛋 白
图3-4 E-Coli在一般情况下,葡萄糖充足,阻抑 物蛋白与lacO结合,抑制转录。
三、乳糖操纵子的诱导表达模型
cAMP 阻抑物蛋白
cAMP 受体蛋 白
图3-5 E-Coli在含有葡萄糖下,进行诱导表达
操纵子学说的意义

何谓操纵子?操纵子学说的重要意义有哪些?
操纵子:是细菌基因表达调控的单元,包括结构基因以及被调节基因产物所识别的存在于DNA分子上的顺式作用元件。
在细菌中,参与某一代谢过程的相关酶的基因成簇排列,其表达受共同的调控序列(调节基因、CAP位点、操纵基因、弱化子等)的调节。
这句是我自己整的,不一定对。
操纵子学说是由法国科学家Francois Jacob,Andre Lwoff 和Jacques Monod提出并得到验证,并于1965年荣获了Nobel 奖。
意义:(1)暗示了“三联体遗传密码”以外的“空间调控密码”的存在;
(2)为分子生物学研究开辟了“中心法则”以外的新领域;
(3)从理论上为遗传学家Barbara Mclintock 早在1947年发现玉米Ac/Ds 遗传“转座因子”的调控假说提供了重要的理论证据。
(4)“调节基因”概念的提出及其普遍性的证实。
操纵子及RNA转录

操纵子(operon)DNA2组中成簇串联组成。
DNA序列。
多种原核基因启及-35一动序列的共有序列在-10区域是TATAAT,又),在TTGACA。
合,或使RNA聚合酶不能沿DNA还有一种特异DNA序列可结合激活蛋白,使转录激活,介导正性调节。
转录(Transcription DNA)UTP)4RNA的过程。
MRNA,通过它携有的密码子到核糖DNA的复制相比,有很多相同或相似之处,亦有其自己的特点。
mRNA。
就是说,以特定的以DNADNA另一条单链叫非模板链。
DNA。
真核细胞的大小亚基是在核中形成的, 在核仁部位rDNA转录出45S rRNA,是rRNA的前体分子,与胞质运来的蛋白质结合,再进行加工,经酶裂解成28S,18S和5.8S的rRNA,而5S rRNA则在核仁外合成28S,5.8S及5S rRNA与蛋白质结合,形成RNP分子团。
为大亚基前体,分散在核仁颗粒区,再加工成熟后,经核孔入胞质为大亚基,18S rRNA也与蛋白质结合,经核孔入胞质为小亚基。
大小亚基在胞质中可解离存在,在需要时也可在>0.001M Mg 存在时,但合成完整单核糖体,才具有合成功能,当Mg4 <0.001M时则又重新解离。
原核细胞的核糖体较小, 沉降系数为70S,相对分子质量为2.5x103 kDa,由50S和30S两个亚基组成; 而真核细胞的核糖体体积较大, 沉降系数是80S,相对分子质量为3.9~4.5x103 kDa, 由60S和40S两个亚基组成。
典型的原核生物大肠杆菌核糖体是由50S大亚基和30S小亚基组成的。
在完整的核糖体中,rRNA约占2/3, 蛋白质约为1/3。
50S大亚基含有34种不同的蛋白质和两种RNA分子,相对分子质量大的rRNA的沉降系数为23S,相对分子质量小的rRNA为5S。
30S小亚基含有21种蛋白质和一个16S的rRNA分子。
碱性纤维素细菌:通过CMC-Na初筛培养基,采用刚果红染色鉴定法筛选纤维素酶产生菌。
3-1.操纵子-原核基因表达调控

• N-end arm of helix reach around to other face (to minor groove)
Hug
l trans-Factor的二聚体或四聚体以对称的方式结合 cis-Factor 表现一种“二重对称性”
dimer as Right Shoes model binding with palindromic Seq. in major groove
IS/I+
iS > I+ iS > iC
等位基因间的显隐关系
I+
OC
Z
Y
A
mRNA
OC失去与阻遏物 特异结合的能力
乳糖
mRNA
I+
O+
等位基因间的显隐关系
OC > O+
cis-dominant
(顺式显性)cis-dominant
The ability of a site (cis-factor) to control adjacent gene irrespective of the presence in the cell of other alleles of the site.
激活RNA 聚合酶启动
•正控制—诱导型操纵子 (多为分解酶类)
w.t. (I+O+P+) 诱导型
iS mut. 超阻突变 (super-repression) 诱导物 不能被激活因子活化
operon 关闭
I+ > iS
e.g. cAMP control (一种通用的控制系统)
E.coli
Glucose Lactose
01乳糖操纵子的调控模式

▪ 遗传学图谱分析指出,Oc突变位于I与Z之 间,所以,lac体系的4个基因的序列为 IOZY。通过这些观察,Jacob和Monod推 断Oc突变代表DNA链上的一个位点或一个
非编码区域,而不是一个基因,因为可编 码的基因具诱导型合成还是 永久型合成,O区域称为操纵基因。
一、 乳糖操纵子的调控模式
▪ 大肠杆菌乳糖操纵子(lactose operon)包括3 个结构基因:Z、Y和A,以及启动子、控制 子和阻遏子等。转录时,RNA聚合酶首先与 启动区(promoter,P)结合,通过操纵区 (operator,O)向右转录。转录从O区的中间开 始,按Z→Y→A方向进行,每次转录出来的 一条mRNA上都带有这3个基因。转录的调控 是启动区和操纵区进行的 .
6. lac操纵子DNA的调控区域--P.O.区
lac P(启动子区)从I基因结束到 mRNA转录起始位点止,共长82bp(-82~ +1)O区就是阻遏物结合区,位于P区后半 部分和转录起始区(-7~+28),该区序列 有对称性,其对称中心点在+11位。P区的 CAMP-CAP结合区(-67~-52)也有对称 性,其对称位点在-60~-59之间。
5. cAMP与代谢物激活蛋白
▪ 当葡萄糖和乳糖同时存在于培养基中时, lac启动子表达受阻,没有β-半乳糖苷酶活 性;当葡萄糖消耗完以后(图中箭头处), 细胞内cAMP浓度增加,β-半乳糖苷酶活性 被诱导,一度停止生长的细胞又恢复分裂。
如果将细菌放在缺乏碳源的培养基中,细 胞内cAMP浓度就很高;若在含葡萄糖的培 养基中培养,细菌中的cAMP浓度就会很低; 如果将细菌置于甘油或乳糖等不进行糖酵 解的碳源培养基中,细菌中cAMP的浓度也 会很高。
②在lac mRNA分子内部,A基因比Z基因更 易受内切酶作用发生降解,因此,在任何时候Z基 因的完整拷贝数要比A基因多。
阿拉伯糖操纵子
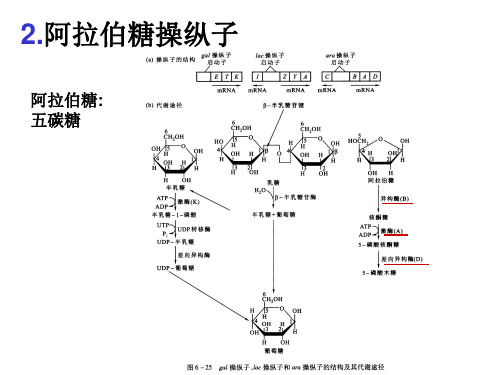
❖araC 基因负控阻遏调节 (自调控)
当细胞内没有AraC蛋白时,由Pc启动子起始araC 基因转录 当AraC的浓度超过40个拷贝/细胞,就与araO1结 合而调控自身的合成,抑制araC基因的转录
III. 营养状况对ara操纵子活性的影响
当AraC蛋白以正调控因子作用时,起始转录还需要 cAMP-CRP的共同参与。
• AraC蛋白的两种形式
AraC蛋白作为PBAD活性正、负调节因子的双 重功能是通过该蛋白的两种异构体来实现
Pr形式
Pi形式
araO2
araI AraC
阻遏 araPBAD
araO2
araI
AraC
激活 araPBAD
阿拉伯糖操纵子的结构
基因的排列序列与代谢途径中酶的作用序列不同
阿拉伯糖操纵子的调控
araBAD和araC基因的转录是分别在两条链上以相反的方向进行的。
AraC, +/-阿拉伯糖:阻遏物 araPC
AraC: 阻遏物, - 阿拉伯糖 激活物, +阿拉伯糖
CAP-cAMP: 激活物
araPBAD
araC 调节基因
EXAMPLES: -araBAD when AraC binds to araI1/araI2 (ligand: arabinose)
-CAP (ligand: cAMP)
3.阻遏蛋白LexA的降解与细菌中的SOS应答
当细菌DNA遭到破坏时,细菌细胞内会启动一 个被称为SOS的诱导型DNA修复系统,参与 SOS DNA修复系统的许多基因分散在染色体的 各个部位,同时受LexA阻遏蛋白的抑制。平时 表达量很低。
操纵子;顺反子

6、多顺反子:多顺反子见于原核生物,意指一个mRNA分子编码多个多肽链。这些多肽链对应的DNA片段则位于同一转录单位内,享用同一对起点和终点。在原核细胞中,通常是几种不同的mRNA连在一起,相互之间由一段短的不编码蛋白质的间隔序列所隔开,这种mRNA叫做多顺反子mRNA。这样的一条mRNA链含有指导合成几种蛋白质的信息。
1、操纵子:(operon):指启动基因(即启动子)、操纵基因和一系列紧密连锁的结构基因的总称,有的操纵子也包含调节基因。在结构基因上游的是启动子序列,能给核糖核酸聚合酶(RNA聚合酶)提供结合位点及引发转录。在启动子附近ቤተ መጻሕፍቲ ባይዱ是一组DNA称为操纵基因。操纵子亦会包含调节基因,如阻遏基因能为调控蛋白质编码,使之与操纵基因结合及阻止转录。调控基因未必是操纵子的一部分,但是位于基因组的某一处。阻遏基因会到达操纵基因阻碍结构基因的转录。操纵子是转录的功能单位,主要见于原核生物的转录调控,如乳糖操纵子、阿拉伯糖操纵子、组氨酸操纵子、色氨酸操纵子等。但真核生物也存在。
就其功能而言,调节基因、操纵基因和启动基因都属于调控基因。
2、结构基因:指编码任何RNA或除了调节因子(regulatory factor)以外的蛋白质的基因。包括编码结构蛋白和酶蛋白的基因,也包括编码阻遏蛋白或激活蛋白的调节基因。它编码的内容呈现广泛的功能和结构,包括结构蛋白、酶类(如催化酶)或不执行调控功能的RNA分子。在真核细胞中,结构基因被内含子和外显子所分隔;而在原核细胞中则是连续的。与调控基因、编码启动子的基因不同,结构基因在蛋白质的翻译中起到实质性的作用。结构基因是一类编码蛋白质或RNA的基因。
操纵子名词解释

操纵子名词解释操纵子名词是指在语法层面上属于一个更大的名词短语的名词。
它不仅能够在句中单独充当名词短语的主语或宾语,还可以在句中作为修饰语、介词宾语等等的成分。
操纵子名词的操纵动作通常是由动词完成的,这个动作直接作用于操纵子名词。
这种语法结构可以使句子更加简洁明了,并且增加句子的表达力。
操纵子名词可以通过两种方式来解释,分别是介词短语和动词的直接宾语。
首先,介词短语可以充当操纵子名词。
例如,在句子“My love for music is strong.”中,"for music"就是一个操纵子名词,且由介词“for”引导。
这个操纵子名词修饰了名词“love”,表达了“对音乐的热爱”之意。
此外,在句子“I have a passion for cooking.”中,“for cooking”也是一个操纵子名词,它修饰了名词“passion”,表达了“对烹饪的热情”之意。
其次,动词的直接宾语可以充当操纵子名词。
例如,在句子“He enjoys playing basketball.”中,“playing basketball”作为动词“enjoys”的直接宾语,可以看作是一个操纵子名词。
它修饰了动词“enjoys”,表达了“喜欢打篮球”之意。
另外,在句子“She hates doing homework.”中,“doing homework”作为动词“hates”的直接宾语,也是一个操纵子名词。
它修饰了动词“hates”,表达了“讨厌做作业”之意。
总结来说,操纵子名词是语法层面上属于一个更大的名词短语的名词,它可以通过介词短语或者动词的直接宾语来进行解释。
通过使用操纵子名词,可以使句子更加简洁明了,并且增加句子的表达力。
分子生物学课程思政教学实践案例——原核生物的基因调控(一)乳糖操纵子

分子生物学课程思政教学实践案例——原核生物的基因调控
(一)乳糖操纵子
李根亮;唐玉莲;李书;孙丽双
【期刊名称】《广东化工》
【年(卷),期】2022(49)3
【摘要】课程思政是思想教育的关键环节,有助提高研学生思想意识形态。
分子生物学是一门感性与理性并存的学科,既有逻辑推理又有人生哲理。
有效地将课程思政与分子生物学融合是提高学生学习能力、思想意识形态和逻辑推理能力的重要手段。
本文以"乳糖操纵子调控机制"教学内容为例,从课堂思政融入教学的整体设计思路、教学内容思政教学点、教学效果和教师感悟等几个方面进行了简要介绍,取得了一定教学效果,希望对其他学科的课程思政融入起到很好的借鉴作用。
【总页数】3页(P193-194)
【关键词】乳糖操纵子;课堂思政;分子生物学;人才培养
【作者】李根亮;唐玉莲;李书;孙丽双
【作者单位】右江民族医学院
【正文语种】中文
【中图分类】G63
【相关文献】
1.课程思政背景下分子生物学课程思政元素教学探索
2.生物化学与分子生物学课程思政实践与思政元素挖掘
3."课程思政"在生物化学与分子生物学教学中的实践与思考
4."课程思政"融入研究生课程口腔分子生物学与实验动物模型的教学实践初探
5.分子生物学融入课程思政的教学研究与实践
因版权原因,仅展示原文概要,查看原文内容请购买。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
乳糖操纵子07生物科学09号李宝青【摘要】本文主要从操纵子的概念着手,简要介绍关于操纵子的基本结构,结构基因群、启动子、操纵基因、调控基因和终止子的结构和基本功能,并介绍了乳糖对乳糖操纵子的诱导调控机制。
【关键词】乳糖操纵子负性调控操纵子学说是关于原核基因结构及其表达调控的学说,由法国巴斯德研究所著名科学家的Monod和Jacob在1961年首先提出[1]。
他们以对乳糖操纵子的研究,通过大量的试验及分析建立了现在已经被人们广泛接受的乳糖操纵子控制模型[2]。
后来人们在大肠杆菌中又陆续发现了色氨酸操纵子、组氨酸操纵子、半乳糖操纵子、阿伯糖操纵子等多种操纵子,从而不断的充实和完善了被誉为生命第三原理的基因调控理论,在这个理论中提出的操纵子概念也被人们普遍接受和证实。
1.乳糖操纵子模型主要内容有:(1)Z、Y、A基因的产物由同一条多顺反子的mRNA分子所编码;(2)该mRNA分子的启动区(P)[3]位于阻遏基因(I)与操纵区(O)之间,不能单独起始半乳糖苷酶和透过酶基因的高效表达;(3)操纵区是DNA上的一小段序列(仅为26bp),是阻遏物的结合位点;(4)当阻遏物与操纵区相结合时,lac mRNA的转录起始受到抑制;(5)诱导物通过与阻遏物结合,改变它的三维构象,使之不能与操纵区相结合,从而激发lac mRNA 的合成。
有人就是说,有阻遏物存在时,操纵区没有被阻遏物占据,所以启动子能够顺利起始mRAN的转录[4]。
2.乳糖操纵子的基本结构2.1结构基因群操纵子中被调控的编码蛋白质的基因称为结构基因。
一个操纵子中含有1个以上的结构基因,多的可达十几个,各结构基因头尾衔接、串连排列,组成结构基因群。
乳糖操纵子含有Z、Y和A共3个结构基因[5]。
Z基因长3510bp,编码含1170个氨基酸、分子量为135000的多肽,以四聚体形式组成有活性的β-半乳糖苷酶,催化乳糖转变为别乳糖(也称异乳糖),再分解为半乳糖[6]和葡萄糖。
Y基因长780bp,编码有260个氨基酸、分子量为30000的半乳糖透过酶,促使环境中的乳糖进人细菌。
A基因长825bp,编码有275个氨基酸、分子量为32000的转乙酞基酶,二聚体活性形式催化半乳糖的乙酞化。
基因Z 的5’侧具有大肠杆菌核糖体识别结合位点特征的Shan-Dagano(SD)序列[7],因而当乳糖操纵子开放时,核糖体能结合在转录产生的mRNA上。
由于Z、Y和A三个基因头尾相接,上一个基因的翻译终止码靠近下一个基因的翻译起始码,因而同一个核糖体能沿此转录生成的多顺反子mRNA移动,在翻译合成了上一个基因编码的蛋白质后、不从mRNA上脱落下来而继续沿mRNA移动合成下一个基因编码的蛋白质,依次合成这个基因群所编码的蛋白质。
2.2启动子[8]启动子是指能被聚合酶识别、结合并启动基因转录的一段DNA序列。
它含有RNA聚合酶特异性结合和转录起始所需的保守序列位点,启动子本身不被转录。
操纵子至少有个启动子,一般在第二个结构基因5‘侧上游,控制整个结构基因一群的转录。
虽然不同的启动子序列有所不同,但比较已经研究过的上百种原核生物的启动子的序列,发现有一些共同的规律,它们一般长40-60bp,含A-T碱基对较多,某些段落是很相似的,这些相似的保守性段落称为共有性序列。
启动子一般可分为识别、结合和起始三个区段。
转录起始第一个碱基(通常标记+1)位置为最常见的是A。
在-10bp附近有TA TAA T一组共有序列,因为这段共有序列是首先发现的,称为Pribnow框[9]。
在-35bp处又有TTGACA一组共有序列。
不同的启动子序列不同,与聚合酶的亲和力不同,启动转录的频率高低不同,即不同的启动子起动基因转录的强弱不同,例如PL、PR、PT7属强启动子,而PLac则是较弱的启动子。
2.3操纵基因操纵基因[10]是指能被调控蛋白特异性结合的一段DNA序列。
操纵基因常与启动子邻近或与启动子序列重叠,当调控蛋白结合在操纵基因序列上,会影响其下游基因转录的强弱。
乳糖操纵子的操纵基因序列位于启动子与被调控的基因之间,部分序列与启动子序列重叠。
研究发现这段双链DNA具有回文样的对称性一级结构,能形成十字形的茎环构造。
不少操纵子都具有类似的对称性序列,可能与特定蛋白质的结合相关。
2.4调控基因调控基因[11]是编码能与操纵序列结合的调控蛋白的基因。
与操纵子结合后能减弱或阻止其调控基因转录的调控蛋白称为阻遏蛋白,其介导的调控方式称为负性调控与操纵子结合后能增强或启动其调控基因转录的调控蛋白称为激活蛋白,所介导的调控方式称为正性调控。
某些特定的物质能与调控蛋白结合,使调控蛋白的空间构象发生变化,从而改变其对基因转录的影响,这些特定物质称为效应物,其中凡能引起诱导发生的分子称为诱导剂,能导致阻遏发生的分子称为阻遏剂或铺助阻遏。
乳糖操纵子中,调控基因Rlac位于Plac邻近,有其自身的启动子和终止子,转录方向和结构基因群的转录方向一致,编码产生由347个氨基酸组成的调控蛋白R。
在环境没有乳糖存在的情况下,R形成分子量为152 000的活性四聚体,能特异性与操纵基因O紧密结合,从而阻止利用乳糖的酶类基因的转录,所以R是乳糖操纵子的阻遏蛋白。
当环境中有足够的乳糖时,乳糖受β一半乳糖晋酶作用转变为异乳糖,异乳糖与R结合,使R的空间构象变化,四聚体解聚成单体,失去与操纵基因特异性紧密结合的能力,从而解除了阻遏蛋白的作用,使其后的基因得以转录合成利用乳糖的酶类。
在这过程中乳糖实际起作用的是别乳糖就是诱导剂,与结合,起到去阻遏作用,诱导了利用乳糖的酶类基因的转录。
许多调控蛋白都是变构蛋白,通过与上述类似的方式与效应物结合改变空间构象,从而改变活性,起到调节基因转录表达的作用2.5终止子终止子[12]是给予聚合酶转录终止信号的序列,它是基因DNA分子中决定转录产物3’-OH端、酶分子停止聚合,释放出已合成RNA分子的位点。
在一个操纵子中至少在结构基因群最后一个基因的后面有一个终止子。
原核生物的终止子结构分为两种主要类型:①内在终止子,是一类不依赖rho因子的终止子(简单终止子);②依赖rho因子的终止子。
不同的终止子的作用也有强弱之分,有的终止子几乎能完全停止转录,有的则只是部分终止转录,一部分聚合酶能越过这类终止序列继续沿移动并转录。
如果一串结构基因群中间有这种弱终止子的存在,则前后转录产物的量会有所不同,这也是终止子调节基因群中不同基因表达产物比例的一种方式。
以上5种元件是每一个操纵子必定含有的其中启动子、操纵子位于紧邻结构基因群的上游,终止子在结构基因群之后,它们都在结构基因的附近,只能对同一条链上DNA的基因表达起调控作用,这种作用在遗传学实验上称为顺式作用,启动子、操纵子和终止子就属于顺式作用元件[13]。
调控基因可以在结构基因群附近,也可以远离结构基因,它是通过其基因产物—调控蛋白来发挥作用的,因而调控基因不仅能对同一条链上DNA的结构基因起表达调控作用,而且能对不在一条链上DNA的结构基因起作用,在遗传学实验上称为反式作用,调控基因就属于反式作用元件[14],其编码产生的调控蛋白称为反式调控因子。
由此,基因表达调控机理的关键在蛋白质与核酸的相互作用上。
3乳糖操纵子的表达调控3.1乳糖的诱导作用乳糖作为原核细胞的常用碳源,是乳糖操纵子控制的细菌自身系列酶的天然底物,对乳糖操纵子具有天然的诱导作用[15-16]。
当细菌在有葡萄糖的培养基中生长时,不能代谢乳糖,因为缺少乳糖代谢的酶。
当生长在没有葡萄糖只有乳糖的培养基中时,代谢乳糖的每两从几个分子迅速增加近千倍即细菌在短时间内合成了能够利用乳糖的一系列酶,具备了利用乳糖作为碳源的能力,在这种培养基上生存下来。
这就是乳糖对操纵子的诱导作用。
细菌获得的这一能力的原因是在乳糖的诱导作用下开启了乳糖操纵子,表达了与代谢乳糖相关的一系列酶所致。
3.2乳糖操纵子的调控机制当培养基中没有乳糖时,存在于操纵子上游的调节基因编码表达的阻遏蛋白结合到操纵子中的操纵基因上,阻止了结构基因的的表达lac基因被调节基因编码表达的阻遏蛋白所阻遏,此时,在细胞中只有几个β-半乳糖苷酶分子。
但将大肠杆菌转到乳糖培养基中时,由于诱导物分子结合在阻遏蛋白的特异部位,引起阻遏蛋白构象变化,不能结合到操纵基因上,是RNA聚合酶能够正常转录存在于从操纵子的结构基因,及操纵子被诱导表达,β-半乳糖苷酶分子数量迅速增加,在这个系统中的诱导物分子不是乳糖本身,而是乳糖的同分异构体—异乳糖,因为乳糖进入大肠杆菌细胞后被转化成了异乳糖。
由于乳糖作为细胞代谢的能源,其浓度是变化的,诱导条件不易掌握[17],其本身作为一种碳源可以被菌体代谢利用,因而对于菌体的生理及代谢也有一定程度的影响。
在实验室中为了准确的得到结果,常常用能被酶识别但不能被分解的半乳糖苷化合物作为诱导剂,如异丙基-β-D-硫代半乳糖苷(简称IPTG)。
IPTG是一种非常高效的乳糖启动子诱导剂,但由于其具有潜在的毒性[18],对菌体生长具有一定的抑制作用,并且价格昂贵,因而不适宜在发酵罐中进行基因工程产品的规模化生产。
IPTG作为诱导剂,其诱导的机制不同于乳糖。
IPTG可以直接进入大肠杆菌细胞内部而发挥诱导作用,且它是一种非代谢性的诱导物,不会被菌体所消耗,只需极少量的存在就能稳定地诱导乳糖启动子的转录;乳糖却需要借助于乳糖透过酶[19]的作用进入细胞,然后经过β-半乳糖苷酶的作用转化为异乳糖(Allolactose)才能起到诱导剂的作用[20-21]。
综上所述,乳糖操纵子属于可诱导操纵子,这类操纵子通常是关闭的,当受效应物作用后诱导转录起始。
这类操纵子使细菌能适应环境的变化,最有效地利用环境能提供的能源物质。
[1]孙乃恩,孙东旭,朱德旭等,《分子遗传学》,南京大学出版社,1990:286[2]Jacob, F. and Monod,J.”Genetic Regulatory Mechanisms in the Synthesi of Proteins.” J.Mol. Biol.1961,3:318~356[3]刘祖洞,江绍慧1遗传学[M]1北京:高等教育出版社,1994:270~2711[4]朱玉贤,李毅,《现代分子生物学》(第二版),高等教育出版社,2002.7,2:201~202[5]张玉静,《分子遗传学》,北京科学出版社,2004.4:231~249[6] 韩贻仁,《分子细胞生物学》,北京科学出版社,2001.3.447~452[7]韦弗(美),《分子生物学》(影印版),北京科学出版社,2008.8.175~190[8] 朱乾浩,《转座子在植物基因分离中的应用研究进展》,《生物工程进展》,1996,V ol.16,NO.2[9]赵亚华,《基础分子生物学教程》,科学出版社,2006:220~221[10] 李汝祺,《中国大百科全书·生物学分册》北京:中国大百科全书出版社,1983:791[11] 广川秀夫(日),胡宝华译,《生物工程名词解释》,化学工业出版社,1991:61[12]褚启人,《遗传的结构与功能》,上海科学技术出版社,1980:317~319[13]张飞雄主编,《普通遗传学》,科学出版社,2004:197~198.[14]张飞雄主编,《普通遗传学》,科学出版社,2004:197~198.[15]郝淑美,王宣军,张秀霞,等.《用乳糖作为诱导剂进行重组蛋白的表达》,中国生物制品学杂志,2005,18(5):409~411.[16]汪建华,熊凌霜,吴军,等人.《重组Shiga2EGF工程菌发酵工艺研究》,中国生化药物杂志,2001,22(6):292~294.[17]Panda A K,Ghorpade A,Mukhopadhyay A,et al.High cell density fermentation of recombinant Vibrio cholerae for the production of B subunit of Eshcherichia coli enterotoxin[J].Biotechnol Bioengineer,1995,45:245~250.[18]金奇.医学分子病毒学.北京:科学出版社,2001.[19]陈亮,任随周,许玫英,孙国萍等人,《乳糖替代IPTG诱导脱色酶TpmD基因在大肠杆菌中的高效表达》,微生物学通报APR 20,2009,36(4):551~556[20]Jobe A,Bourgeois c repressor-operator interaction:Ⅵ.The natural inducer of the lac operon.J Mol Biol,1972,69(3):397~404.[21]Muller-Hill B,Rickenberg HV,Wallenfels K.Specificity of the induction of the enzymes of the lac operon in Es-cherichia coli.J Mol Biol,1964,10:303~318.。