生化课件16转录
合集下载
转录PPT课件
(GTP)
转录起始复合物: RNApol (2) - DNA - pppGpN- OH 3
(4)合成第一个磷酸二酯键后, σ亚基脱落。
第32页/共122页
σ亚基辨认-35区
(结合松弛)
RNA pol 全酶移向-10区 (结合紧密)
合成第一个磷酸二酯键 5’-pppGpN-OH-3’
转录起始复合物
σ亚基脱落
RNA pol(αββ′σ ) —DNA—pppGpN-OH
转录起始不需引物!
第33页/共122页
16/50
二、 原核生物的转录延长时蛋白质的翻译也 同时进行
1. 亚基脱落,RNA–pol聚合酶核心酶变构, 与模板结合松弛,沿着DNA模板前移;
2. 在核心酶作用下,按模板的指引,NTP不 断聚合,RNA链不断延长。(A U) (NMP) n + NTP (NMP) n+1 + PPi
功能atp酶活性解螺旋酶活性能控制转录终止的蛋白质一依赖rho因子的转录终止第40页共122页结合后因子和rna聚合酶都可能収生构象变化从而使rna聚合酶停顿解螺旋酶的活性使rnadna杂化链解链转录的rna释放出来而终止转录第41页共122页模板模板dnadna链在接近转链在接近转录终止点处存在相连录终止点处存在相连的富含的富含gcgc的回文结构的回文结构使使rnarna转录产物形成转录产物形成収夹形的二级结构収夹形的二级结构其后又有一段其后又有一段寡聚寡聚uu
(1)RNA聚合酶只有 5′→3′的聚合酶活性,无5′ 或3′外切酶的活性。RNA聚合酶缺乏 3′-外切酶的 活性是转录忠实性不如复制的主要原因;
(2)RNA聚合酶本身就能够促进DNA双链解链; (3)RNA聚合酶催化转录的从头合成,不需要引物; (4) RNA聚合酶的底物是NTP,而不是dNTP,由UTP
转录起始复合物: RNApol (2) - DNA - pppGpN- OH 3
(4)合成第一个磷酸二酯键后, σ亚基脱落。
第32页/共122页
σ亚基辨认-35区
(结合松弛)
RNA pol 全酶移向-10区 (结合紧密)
合成第一个磷酸二酯键 5’-pppGpN-OH-3’
转录起始复合物
σ亚基脱落
RNA pol(αββ′σ ) —DNA—pppGpN-OH
转录起始不需引物!
第33页/共122页
16/50
二、 原核生物的转录延长时蛋白质的翻译也 同时进行
1. 亚基脱落,RNA–pol聚合酶核心酶变构, 与模板结合松弛,沿着DNA模板前移;
2. 在核心酶作用下,按模板的指引,NTP不 断聚合,RNA链不断延长。(A U) (NMP) n + NTP (NMP) n+1 + PPi
功能atp酶活性解螺旋酶活性能控制转录终止的蛋白质一依赖rho因子的转录终止第40页共122页结合后因子和rna聚合酶都可能収生构象变化从而使rna聚合酶停顿解螺旋酶的活性使rnadna杂化链解链转录的rna释放出来而终止转录第41页共122页模板模板dnadna链在接近转链在接近转录终止点处存在相连录终止点处存在相连的富含的富含gcgc的回文结构的回文结构使使rnarna转录产物形成转录产物形成収夹形的二级结构収夹形的二级结构其后又有一段其后又有一段寡聚寡聚uu
(1)RNA聚合酶只有 5′→3′的聚合酶活性,无5′ 或3′外切酶的活性。RNA聚合酶缺乏 3′-外切酶的 活性是转录忠实性不如复制的主要原因;
(2)RNA聚合酶本身就能够促进DNA双链解链; (3)RNA聚合酶催化转录的从头合成,不需要引物; (4) RNA聚合酶的底物是NTP,而不是dNTP,由UTP
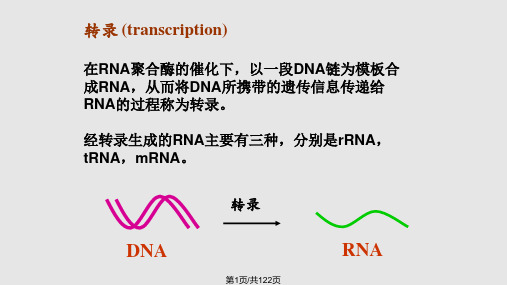
生化课件第十六章 RNA的生物合成

复制和转录的区别
模板 原料 酶 产物
配对 引物
复制
转录
两股链均复制 模板链转录(不对称转录)
dNTP
NTP
DNA聚合酶
RNA聚合酶(RNA-pol)
子代双链DNA mRNA,tRNA,rRNA (半保留复制)
A-T,G-C 需要(RNA)
A-U,T-A,G-C 不需要
目录
第一节 模板和酶
Templates and Enzymes
可以增强对各种损伤的抵抗力. 如热、乙醇、亚砷酸钠、 缺血的预处理.
目录
三、模板上酶的辨认、结合
原核生物一个转录区段可视为一个转录单位,称 为 操 纵 子 (operon) , 包 括 若 干 个 结 构 基 因 及 其 上 游 (upstream)的调控序列。
调控序列
结构基因
5
3
3
RNA-pol
目录
5′···GCAGTACATGTC ···3′ 3′··· c g t c a t g t a c a g ···5′
转录 5′···GCAGUACAUGUC ···3′
翻译
编码链 模板链
mRNA
N······Ala ·Val ·His ·Val ······C 蛋白质
转录生成的RNA链与编码链的碱基序列相似,以U取代T
目录
RNA聚合 酶保护法
目录
第二节 转录过程
The Process of Transcription
目录
大肠杆菌转录过程包括转录起始、延长、终止三步。 真核生物的转录,除延长过程相似外,起始、终止 都与原核生物有较多的不同。
目录
一、原核生物的转录过程
(一)转录起始
转录 ppt课件

启动子的一致序列是综合统计了多种基因的启动子序
列以后得出的结果,迄今为止,在E.coli中还没有发
现哪一个基因的启动子序列与一致序列完全一致。一
个基因的启动子序列与一致序列越相近,则该启动子
的启动效率就越高。不同基因在启动子序列上的差异,
是基因表达调控的一种重要途径。
ppt课件
18
3.2.3 转录的起始
ppt课件
13
因子与其相应的启动子
因子 基因 功能
-35区
间隔
-10区
70 rpoD 广泛
TTGACA
15-17bp TATAAT
32 rpoH 热休克 TCTCNCCCTTGAA 13-15bp CCCCATNTA
54 rpoN 氮代谢
CTGGNA
6bp
TTGCA
ppt课件
14
3.2.2 启动子的识别序列
ppt课件
12
不同的σ因子可以识别不同的启动子。如E.coli的
一般基因由σ70(Mr为70×103)识别, 枯草芽孢杆菌σ因 子的主要类型为σ43。识别热休克应激蛋白基因启动
子的为σ32,温度升高时,基因rpoH的产物σ32浓度升
高,温度回降时,σ32浓度降低。可以假设,σ70与σ32 通过竞争已有的核心酶,调控蛋白质合成的起始。诱 导σ32产生的基本信号是因温度升高引起的未折叠蛋 白的聚集。在固氮菌中识别固氮酶相关基因启动子的
为σ54,当培养基中缺乏氮时,E.coli中会有少量σ54存
在,在这种情况下,基因会转向利用其它可替代的N 源。
ω亚基由基因rpoZ编码,Mr为1.10×104,曾长期
被忽略,甚至许多人不把它作为聚合酶的组分。然而, 现在已经肯定,ω亚基是嗜热水生菌RNA pol必不可 少的组分,也是体外变性的RNA pol成功复性所必需 的,它与β亚基一起构成催化中心,稳定其与β' 亚基 的结合
列以后得出的结果,迄今为止,在E.coli中还没有发
现哪一个基因的启动子序列与一致序列完全一致。一
个基因的启动子序列与一致序列越相近,则该启动子
的启动效率就越高。不同基因在启动子序列上的差异,
是基因表达调控的一种重要途径。
ppt课件
18
3.2.3 转录的起始
ppt课件
13
因子与其相应的启动子
因子 基因 功能
-35区
间隔
-10区
70 rpoD 广泛
TTGACA
15-17bp TATAAT
32 rpoH 热休克 TCTCNCCCTTGAA 13-15bp CCCCATNTA
54 rpoN 氮代谢
CTGGNA
6bp
TTGCA
ppt课件
14
3.2.2 启动子的识别序列
ppt课件
12
不同的σ因子可以识别不同的启动子。如E.coli的
一般基因由σ70(Mr为70×103)识别, 枯草芽孢杆菌σ因 子的主要类型为σ43。识别热休克应激蛋白基因启动
子的为σ32,温度升高时,基因rpoH的产物σ32浓度升
高,温度回降时,σ32浓度降低。可以假设,σ70与σ32 通过竞争已有的核心酶,调控蛋白质合成的起始。诱 导σ32产生的基本信号是因温度升高引起的未折叠蛋 白的聚集。在固氮菌中识别固氮酶相关基因启动子的
为σ54,当培养基中缺乏氮时,E.coli中会有少量σ54存
在,在这种情况下,基因会转向利用其它可替代的N 源。
ω亚基由基因rpoZ编码,Mr为1.10×104,曾长期
被忽略,甚至许多人不把它作为聚合酶的组分。然而, 现在已经肯定,ω亚基是嗜热水生菌RNA pol必不可 少的组分,也是体外变性的RNA pol成功复性所必需 的,它与β亚基一起构成催化中心,稳定其与β' 亚基 的结合
生物化学 - 转录

杂交链中A- U间氢键相对较弱,新生RNA易从模板上脱落,不需ρ因子即 可终止。
不 需 因 子 的 终 止
ρ
发夹结构 寡聚U片段
B、需ρ因子终止 ρ因子是一个6聚体蛋白,其功能: 1)终止因子; 2)NTPase活性; 3)解旋酶活性。 过程: 1)在RNA存在下,水解NTP产能,使ρ因子与 RNApol结合; 2)解旋酶活性使RNA:DNA杂合链拆开,释放RNA,酶和ρ因子一 起从DNA上脱落下来。
ATP
需 因 子 终 止
ρ
(二) 真核细胞的转录 特点: --因有核膜相隔,没有转录与翻译同步的现象。 --一个基因上可同时进行多个转录过程,产生多条RNA链。 --蛋白质编码基因多是不连续的,编码部分(外显子)被不编码基因(内 含子)隔断。
五、转录产物的加工 在专一酶作用下,切除多余部分或修饰,才成为“成熟的”RNA。 • 细菌中合成的mRNA大多不需要加工; • 真核细胞合成的mRNA需要加工。 涉及:(1) 5’-端的帽化(甲基化的鸟苷酸) (2) 3’-端的聚腺苷酸化(polyA) (3) hnRNA、snRNA的剪接 (4) 碱基修饰
帽子结构
鸡卵清蛋白基因
hnRNA
首、尾修饰
hnRNA剪接
成熟的mRNA
鸡 卵 清 蛋 白 基 因 及 其 转 录 、 转 录 后 修 饰
RNA聚合酶
基因起点
5-pppG -OH + NTP 5-pppGpN - OH 3 + ppi
DNA
转录起始复合物:
RNApol (2) - DNA - pppGpN- OH 3
2、延伸 A、 合成开始后,σ亚基释放,然后都由核心酶催化。 B、核心酶沿模板移动,选择NTP,暂时形成DNA-RNA杂交链。
浙江农林大学生物化学--转录PPT课件

❖ β亚基与底物(NTP)及新生RNA链结合; ❖ β′亚基与模板DNA结合; ❖ β和β′亚基组成酶的活性中心,通过DNA的磷酸基团与核心
酶的碱性基团间的非特异性吸附作用,核心酶能与模板DNA 非特异性松驰结合; ❖ σ亚基的功能是识别启动子,辩认转录起始点,但不能单独 与DNA模板结合,当它与核心酶结合时,可引起酶构象的改 变,从而改变核心酶与DNA结合的性质,使全酶对转录起始 点的亲和力比其他部位高4个数量级,在转录延长阶段,σ亚 基与核心酶分离,仅由核心酶参与延长过程。
-
4
第二节 转录所需的条件
一、RNA聚合酶, 二、启动子(-10序列和-35序列), 三、各种转录因子(TFA、TFB、TFD)。 四、四种NTP。
-
5
一、RNA聚合酶
❖ 转录酶(transcriptase)是依赖DNA的RNA聚合酶 (DNA dependent RNA polymerase,DDRP), 亦称为DNA指导的RNA聚合酶(DNA directed RNA polymerase),简称为RNA聚合酶(RNA pol)。 它以DNA为模板催化RNA的合成。
-
14
第四节 真核生物的转录
一、真核生物与原核生物转录的区别 1、转录与翻译部位不同 2、RNA聚合酶不同 3、启动子不二、真核生物的转录过程
一)转录的起始
1、RNA聚合酶I催化的转录起始RNA聚合酶I催化前 rRNA(40S RNA)的合成。前rRNA基因转录起始 点上游有两个顺式作用元件,一个是跨越起始点的 核心元件,另一个在–100bp处有上游调控元件( UCE)。RNA聚合酶I催化的转录需要2种转录因子 ,分别称为上游结合因子(UBF)和选择性因子1 (SL1)。SL1含有4个亚基,一个是TATA盒结合蛋 白(TBP),另3个是TBP相关因子(TAF)。UBF 与DNA结合令模板DNA发生弯曲,使相距上百bp的 UCE和核心元件靠拢,接着SL1和pol I相继结合到 UBF-DNA复合物上,完成起始复合物的组建,开始 转录 。
酶的碱性基团间的非特异性吸附作用,核心酶能与模板DNA 非特异性松驰结合; ❖ σ亚基的功能是识别启动子,辩认转录起始点,但不能单独 与DNA模板结合,当它与核心酶结合时,可引起酶构象的改 变,从而改变核心酶与DNA结合的性质,使全酶对转录起始 点的亲和力比其他部位高4个数量级,在转录延长阶段,σ亚 基与核心酶分离,仅由核心酶参与延长过程。
-
4
第二节 转录所需的条件
一、RNA聚合酶, 二、启动子(-10序列和-35序列), 三、各种转录因子(TFA、TFB、TFD)。 四、四种NTP。
-
5
一、RNA聚合酶
❖ 转录酶(transcriptase)是依赖DNA的RNA聚合酶 (DNA dependent RNA polymerase,DDRP), 亦称为DNA指导的RNA聚合酶(DNA directed RNA polymerase),简称为RNA聚合酶(RNA pol)。 它以DNA为模板催化RNA的合成。
-
14
第四节 真核生物的转录
一、真核生物与原核生物转录的区别 1、转录与翻译部位不同 2、RNA聚合酶不同 3、启动子不二、真核生物的转录过程
一)转录的起始
1、RNA聚合酶I催化的转录起始RNA聚合酶I催化前 rRNA(40S RNA)的合成。前rRNA基因转录起始 点上游有两个顺式作用元件,一个是跨越起始点的 核心元件,另一个在–100bp处有上游调控元件( UCE)。RNA聚合酶I催化的转录需要2种转录因子 ,分别称为上游结合因子(UBF)和选择性因子1 (SL1)。SL1含有4个亚基,一个是TATA盒结合蛋 白(TBP),另3个是TBP相关因子(TAF)。UBF 与DNA结合令模板DNA发生弯曲,使相距上百bp的 UCE和核心元件靠拢,接着SL1和pol I相继结合到 UBF-DNA复合物上,完成起始复合物的组建,开始 转录 。
转录ppt(共66张PPT)

RNA 聚合酶
Ⅰ
Ⅱ
Ⅲ
转录
产物
加工
45SrRNA
18SrRNA 28SrRNA
hnRNA tRNA 5SRNA
snRNA
mRNA
利福平
抑制原核生物的RNA聚合酶
鹅膏蕈碱
抑制真核生物的RNA聚合酶
利福平 主要用于治疗结核病、麻风病等
鹅膏蕈碱 鬼笔鹅膏
三、真核生物的转录产物为单顺
反子
与原核生物不同,真核生物一个转 录单位仅生成一个mRNA分子,经翻 译生成一条多肽链
(三)都形成3’, 5’-磷酸二酯键
二、复制与转录的不同点
模板
原料
酶
校读
复制 两条链 dNTP DNA聚合酶
有
转录 一条链
NTP RNA聚合酶
无
配对
产物
引物
复制
A=T GC
子代双
链DNA
需要
转录
A=U T=A
GC
mRNA tRNA rRNA
不需要
第一节 原核生物的转录
一、转录模板 二、RNA聚合酶 三、模板和酶的辨认结合
第 11 章
RNA的生物合成(转录)
DNA DNA DNA
RNA
蛋白质
中心法则
目录
前 言 复制与转录的异同 第1节 原核生物的转录
第2节 真核生物的转录特点
第3节 真核生物转录后加工
前言 复制与转录的异同
一、相同点 二、不同点
一、复制与转录的相同点
(一)服从碱基配对规则
(二)合成方向都是5’-3’
DNA-dependent RNA polymerase (二)合成方向都是5’-3’ (四)腺嘌呤脱氨成为次黄嘌呤 σ亚基从三元起始复合物上脱落后,核心酶的构象随之发生改变,并沿着模板链的3’→5’方向滑行,进入延长阶段 第1节 原核生物的转录 原核生物的RNA聚合酶仅1种,合成全部RNA 主要用于治疗结核病、麻风病等 真核生物rRNA转录后加工 α2 β β’ ω σ称为全酶
《转录和翻译》课件

转录和翻译的研究
04
进展
转录和翻译的最新研究成果
基因表达调控机制
随着基因组学和分子生物学的发展,科学家们对基因表达 调控机制有了更深入的了解,特别是转录和翻译过程中的 调控机制。
非编码RNA的作用
近年来,越来越多的研究表明,非编码RNA在转录和翻译 过程中发挥着重要作用,为理解基因表达调控提供了新的 视角。
表观遗传学的影响
表观遗传学研究显示,DNA甲基化、组蛋白修饰等表观遗 传学机制对转录和翻译过程具有重要影响,进一步揭示了 基因表达调控的复杂性。
转录和翻译的研究前景
01
人工智能在基因表达调控中的应用
随着人工智能技术的发展,利用人工智能方法对基因表达调控进行预测
、模拟和优化成为研究热点,有望为疾病诊断和治疗提供新的手段。
《转录和翻译》课件
目录
• 转录 • 翻译 • 转录与翻译的相互关系 • 转录和翻译的研究进展
转录
01
转录的定义
01
转录转录是指以DNA为模板合成 RNA的过程。
02
转录转录是基因表达的第一步, 它发生在细胞核中,通过RNA聚 合酶的作用,以DNA为模板合成 RNA分子。
转录的过程
转录转录过程包括启动、延伸和终止三个阶段。
转录在转录过程中,RNA聚合酶首先识别并结合到DNA模板链上的启动子区域 ,然后开始合成RNA链。在延伸阶段,RNA聚合酶以5'-3'方向持续合成RNA, 直到遇到终止信号。最后,RNA聚合酶从DNA模板上释放出来,转录终止。
转录的调控
转录转录调控是通过各种机制对转录过程进行调节和控制的 。
转录转录调控可以发生在转录的各个阶段,包括启动子区域 的调节、RNA聚合酶的调节以及转录过程中的其他调控因子 。这些调控机制可以影响基因的表达水平,从而影响细胞的 功能和行为。
(精选)《转录及转录后加工》PPT课件

10
RNA聚合酶——
二、真核生物的RNA聚合酶
真核生物的RNA聚合酶
种类
Ⅰ
Ⅱ
Ⅲ
定位 转录产物
核仁 45s-rRNA
对鹅膏蕈碱反应 耐受
核质 hnRNA U1-13snRNA (U6除外) 极敏感
核质
5s-rRNA,tRNA, U6snRNA, 非UsnRNA 中度敏感
11
转录模板
• DNA分子上转录出RNA的区段,称为结构基因 (structural gene)。
T T T A C A…N17…T A T G T T · N6 · A…
T T G A T A…N16…T A T A A T · N7 · A…
C T G A C G…N18…T A C T G T · N6 · A…
TTGACA
38 36 29 37 37 28
TATAAT
40 25 30 41 29 44
16
调控序列
结构基因
5 3
RNA-pol
3 5
RNA聚合酶结合模板DNA的部位, 称为启动子(promoter)。
17
RNA聚合 酶保护法
18
RNA聚合酶保护区 结构基因
5
3
3
5
5
3
-50 -40 -30 -20 -10 1 10
3
5
-35 区
TTGACA AA C T G T
RNA-pol辨认位点 (recognition site)
7
RNA聚合酶——
大肠杆菌RNA聚合酶组分
亚基
分子量
36512 150618 155613 70263
功能
决定哪些基因被转录 催化功能
RNA聚合酶——
二、真核生物的RNA聚合酶
真核生物的RNA聚合酶
种类
Ⅰ
Ⅱ
Ⅲ
定位 转录产物
核仁 45s-rRNA
对鹅膏蕈碱反应 耐受
核质 hnRNA U1-13snRNA (U6除外) 极敏感
核质
5s-rRNA,tRNA, U6snRNA, 非UsnRNA 中度敏感
11
转录模板
• DNA分子上转录出RNA的区段,称为结构基因 (structural gene)。
T T T A C A…N17…T A T G T T · N6 · A…
T T G A T A…N16…T A T A A T · N7 · A…
C T G A C G…N18…T A C T G T · N6 · A…
TTGACA
38 36 29 37 37 28
TATAAT
40 25 30 41 29 44
16
调控序列
结构基因
5 3
RNA-pol
3 5
RNA聚合酶结合模板DNA的部位, 称为启动子(promoter)。
17
RNA聚合 酶保护法
18
RNA聚合酶保护区 结构基因
5
3
3
5
5
3
-50 -40 -30 -20 -10 1 10
3
5
-35 区
TTGACA AA C T G T
RNA-pol辨认位点 (recognition site)
7
RNA聚合酶——
大肠杆菌RNA聚合酶组分
亚基
分子量
36512 150618 155613 70263
功能
决定哪些基因被转录 催化功能
转录的基本过程课件.ppt

精
10
精
11
RNA转录的基本过程
精
1
RNA链合成的特点
• RNA是按5’→3’方向合成 • 以DNA双链中的反义链(模板链)为模板 • 以4种三磷酸核苷(NTPs)为原料 • 根据碱基配对原则 • 各核苷酸间通过形成磷酸二酯键相连 • 不需要引物 • 合成的RNA带有于DNA编码链相同的序列
精
2转录的基本过程精8三、转录延伸
• 转录延伸:RNA聚合 酶释放σ 因子离开启 动子,核心酶沿模板 DNA链移动并使新生 RNA链不断伸长的过 程。
精
9
四、转录终止
• 转录终止就是当RNA链延伸到转录终止位 点时,RNA聚合酶不再形成新的磷酸二酯 键,RNA-DNA杂合物分离,转录泡瓦解, DNA恢复成双链状态,而RNA聚合酶和RNA 链都被从模板上释放出来的过程。
1rna链合成的特点链合成的特点rna是按53方向合成以dna双链中的反义链模板链为模板以4种三磷酸核苷ntps为原料根据碱基配对原则各核苷酸间通过形成磷酸二酯键相连不需要引物合成的rna带有于dna编码链相同的序列转录的基本过程转录的基本过程模板识别转录起始转录延伸转录终止一模板识别一模板识别模板识别阶段主要指rna聚合酶与启动子dna双链相互作用并与之相结合的过程
上第一个核苷酸键的 产生。
精
7
通过启动子的时间代表一个启动子的强弱 • 转录起始后直到形成9个核苷酸短链的过程是通
过启动子阶段,此时RNA聚合酶一直处于启动子 区,新生的RNA链与DNA模板链的结合不够牢固, 很容易从DNA链上掉下来并导致转录重新开始。
• 一旦RNA聚合酶成功地合成9个以上核苷酸并离开 启动子区,转录就进入正常的延伸阶段。
• 模板识别 •↓ • 转录起始 •↓ • 转录延伸 •↓ • 转录终止
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
TF II E
(6)TF II H 和TF II J加入复合物
(7)TF II H
有多种酶活性,包括ATP酶、解旋酶、和可使Pol II 的
CTD 磷酸化的激酶活性。
Pol II 的CTD 磷酸化
, TF II 在PolⅡ离开启动 子前释放,形成适于 延伸的构象
Pol II 离开启动子区,
进入延伸阶段
用。
TF II B
(4)与RNA聚合酶与TFIIF相连的复合体结合
TF II F
Pol II
TF II F 结合Pol II并带向启动子; 两个亚基:
RAP74(ATP依赖性解旋酶),可能参与DNA 双链的溶解 RAP30(与细菌因子有同源性),与RNA 聚合酶Ⅱ紧密 结合
(5)TF II E 扩大DNA覆盖区至+30
RNA聚合酶Ⅱ起始复合物的组装 启动子TATA盒+ TFⅡD
+ TFⅡA + TFⅡB +(RNA聚合酶+ TFⅡF)复合物 +TFⅡE 、TFⅡJ +TFⅡH(解旋酶、蛋白激酶)
DNA解旋、RNA聚合酶Ⅱ的 CTD磷酸化
polⅡ从转录因子中释放出来 从起始点向下游移动
转录的终止 (1)终止位点:
(以E.coli为例,五聚体)
核心酶 (core enzyme) 全酶 (holoenzyme) 利福平
三、RNA聚合酶与启动子
1、转录是不连续、分区段进行的 2、每一个转录区段可视为一个转录单位,包括 若干个结构基因及其上游(upstream)的调控序列
结合部位: RNA聚合酶的结合的部位,其长度为7bp,
中心部位在–10bp处,一致性序列为TATAAT序列,故称 之为 TATA box (Pribnow box)
识别部位和结合部位都富含AT碱基,因此双链DNA易发 生解链,有利于RNA聚合酶的结合
第二节
转录过程
The Process of Transcription
D
非编码区
2. 外显子(exon)和内含子(intron)
• 外显子 在断裂基因及其初级转录产物上出现,
并表达为成熟RNA的核酸序列
•内含子 隔断基因的线
性表达而在剪接过程中 被除去的核酸序列
鸡
鸡卵清蛋白
卵
基因
清
蛋
白
基
hnRNA
因
及
首、尾修饰
其
转
录
、
hnRNA剪接
转
录
后
成熟的mRNA
修
饰
3. mRNA的剪接 *
转录空泡
5’ 3’
3’
3’
5’
5’
RNA-pol (核心酶)-DNA -RNA
(四)转录终止 原核生物存在两类终止方式
1.非依赖Rho (ρ)的转录终止子
序列特征:回文序列富含GC,且在下游为连续的A
2.依赖Rho的转录终止子
序列特征:回文序列GC含量相对较少,下游序列无明显 特征
5’
3’
3’
5’
依赖 Rho因子的转录终止
◆Rho因子是rho基因的产物,广泛存在于原核和真 核细胞中,由6个亚基组成,分子量300KD。Rho因 子结合在新生的RNA链上,由5’端向3’端利用ATP 水解供能快速移动 ◆ Rho因子有ATP酶的活性和解螺旋酶的活性
DNA ρ因子
RNA聚合酶 RNA
第三节 真核生物RNA的生物合成
反式作用因子中,直接或间接结合RNA聚 合酶的,则称为转录因子(transcriptional factors, TF)
RNApol II 的转录因子
TF II D TBP(TATA盒结合蛋白) + TAFs(TBP协同因子 )
—— 结合在DNA小沟(其他DNA结合蛋白为大沟),识 别和结合核心启动子(TATA盒和Inr)
启动子
调控序列
5 3
RNA-pol
结构基因
3 5
目录
启动子
RNA聚合酶结合模板DNA的部位,称为启 动子(promoter)
RNA聚合酶 保护法
RNA聚合酶保护区 结构基因
5
3
3
5
5
3
-50 -40 -30 -20 -10 1 10
3
5
-35 区 TTGACA AACTGT
原核生物启动子保守序列
目前还不清楚RNA聚合酶Ⅱ基因的精确终止位点 (2)AAUAAA序列:
初始转录物3’末端的保守序列
对于转录产物的准确切割及加poly(A)是必须的
起始
延伸
5’ cap 5’ cap
AAUAAA AAUAAA
RNA内切酶 AAUAAA识别因子
Poly(A)聚合酶 An
Roger D. Kornberg
TATA
-40 -30 -20 -10
+10 +20
TBP的作用----定位因子 a、在TATA框处与DNA
结合
b、是所有三种RNA聚合 酶转录起始所需的因 子
TBP的作用机制: a 、两个结构域形成一个完全二元对称的马鞍型 结构,与DNA小沟有效结合
b、TBP 与DNA 结合,使DNA 弯曲了约80, TATA 盒向大沟弯曲,拓宽了小沟。
(2) TFIIA
含有至少3个亚基 与TFIID结合,稳定TFIID-DNA复合体;可能通过解除
TAFs的抑制而激活TBP
TF II A
(3) TFIIB 覆盖靠近起始点的启动位置,C端与TFIID和DNA的复合
物结合,N-端与TFⅡF协同作用募集RNA聚合酶II
TFIIB与TFIID结合,并为RNA聚合酶结合起一个桥梁作
结合,这种构象适于转录的起始 (2)CTD磷酸化可使RNA聚合酶II与DNA的
结合变得松弛,形成适于延伸的构象
RNA聚合酶II自身不能起始转录,需要依靠转
录因子的协助
转录因子
能直接、间接辨认和结合转录上游区段 DNA的蛋白质,现已发现数百种,统称为反 式作用因子(trans-acting factors)
目录
RNA聚合酶Ⅱ 与转录因子 羧基末端结构域(CTD): 1、位置:RNA聚合酶Ⅱ最大亚基羧基末端
2、结构特点: (1)具有7个氨基酸(Tyr-Ser-Pro-Thr-Ser -Pro-Ser)
的重复序列(酵母:重复26次;哺乳类:52次 )
(2)多个磷酸化位点:Ser、Thr
CTD磷酸化对调控基因转录有重要作用: (1)CTD去磷酸化,RNA聚合酶II易与DNA
4. 亚基脱落
(二)转录延长
1. 亚基脱落,核心酶变构,与模板结合松弛 ,沿着DNA模板前移
2. 在核心酶作用下,NTP不断聚合,RNA链 不断延长
(NMP) n + NTP (NMP) n+1 + PPi
转录空泡(transcription bubble):也称转录复合物,是由 RNA聚合酶的核心酶、DNA模板和转录产物RNA三者结合在 一起的复合体
目录
5′···GCAGTACATGTC ···3′ 编码链 3′··· c g t g a t g t a c a g ···5′ 模板链
转录 5′···GCAGUACAUGUC ···3′
翻译
mRNA
N······Ala ·Val ·His ·Val ······C蛋白质
目录
结构基因
5
编码链
3
模板链
转录方向
转录方向
模板链
3
编码链
5
不对称转录
(asymmetric transcription)
• 在DNA分子双链上某一特定区段,一股链 用作模板指引转录,另一股链不转录
• 模板链不总是在同一条单链上
二、RNA聚合酶
1、合成机制与DNA-pol相似 2、不需要引物
RNA聚合酶
(二)原核生物的RNA聚合酶
5 m7GpppGp…
帽子的作用:
增强mRNA的稳定性,防止被核酸酶水 解
蛋白质合成时:促进核糖体与mRNA的 结合,启动蛋白质的合成
(二)3’-末端:polyA(尾)
尾巴的作用: 参与mRNA从核内向胞质的定向转移 防止外切酶的水解,增强mRNA的稳定性
(三)mRNA的剪接 hnRNA
• 核内的初级转录产物。称为杂化核RNA (hetero-nuclear RNA, hnRNA)或核内不均一 RNA
TF II E —— 扩大DNA覆盖区至+30
TF II H 和TF II J —— H有激酶活性, 使Pol II 的CTD 磷酸化 ,PolⅡ 离开启动子区
RNA PolⅡ基因转录的过程 1、转录起始
(1) TFIID:
TBP(TATA盒结合蛋白)
因子 )
+ TAFs(TBP协同
TBP TAFs
回文序列
终止子: DNA模板上能够在转录即将结束时,提供终止信号的序 列,称为终止子(terminator)。通常为一段被间隔开的回文序列 ,存在于RNA pol已经转录过的序列中
RNA-pol
5
3
3
5
5´pppG
茎环结构使转录终止的机理 • 使RNA聚合酶变构,阻碍前移;
• 使杂交双链易于解离,转录复合物解体,RNA 产物释放
TF II A —— 含数个亚基,可能通过解除TAFs的抑制而激活TBP
TF II B —— 覆盖靠近起始点的启动位置,C端与TFIID和DNA 的复合物结合,N-端与TFⅡF协同作用募集RNA聚 合酶II
TF II F ——结合Pol II并带向启动子;RAP74(ATP依赖性解 旋酶),RAP30(与细菌因子有同源性)
(6)TF II H 和TF II J加入复合物
(7)TF II H
有多种酶活性,包括ATP酶、解旋酶、和可使Pol II 的
CTD 磷酸化的激酶活性。
Pol II 的CTD 磷酸化
, TF II 在PolⅡ离开启动 子前释放,形成适于 延伸的构象
Pol II 离开启动子区,
进入延伸阶段
用。
TF II B
(4)与RNA聚合酶与TFIIF相连的复合体结合
TF II F
Pol II
TF II F 结合Pol II并带向启动子; 两个亚基:
RAP74(ATP依赖性解旋酶),可能参与DNA 双链的溶解 RAP30(与细菌因子有同源性),与RNA 聚合酶Ⅱ紧密 结合
(5)TF II E 扩大DNA覆盖区至+30
RNA聚合酶Ⅱ起始复合物的组装 启动子TATA盒+ TFⅡD
+ TFⅡA + TFⅡB +(RNA聚合酶+ TFⅡF)复合物 +TFⅡE 、TFⅡJ +TFⅡH(解旋酶、蛋白激酶)
DNA解旋、RNA聚合酶Ⅱ的 CTD磷酸化
polⅡ从转录因子中释放出来 从起始点向下游移动
转录的终止 (1)终止位点:
(以E.coli为例,五聚体)
核心酶 (core enzyme) 全酶 (holoenzyme) 利福平
三、RNA聚合酶与启动子
1、转录是不连续、分区段进行的 2、每一个转录区段可视为一个转录单位,包括 若干个结构基因及其上游(upstream)的调控序列
结合部位: RNA聚合酶的结合的部位,其长度为7bp,
中心部位在–10bp处,一致性序列为TATAAT序列,故称 之为 TATA box (Pribnow box)
识别部位和结合部位都富含AT碱基,因此双链DNA易发 生解链,有利于RNA聚合酶的结合
第二节
转录过程
The Process of Transcription
D
非编码区
2. 外显子(exon)和内含子(intron)
• 外显子 在断裂基因及其初级转录产物上出现,
并表达为成熟RNA的核酸序列
•内含子 隔断基因的线
性表达而在剪接过程中 被除去的核酸序列
鸡
鸡卵清蛋白
卵
基因
清
蛋
白
基
hnRNA
因
及
首、尾修饰
其
转
录
、
hnRNA剪接
转
录
后
成熟的mRNA
修
饰
3. mRNA的剪接 *
转录空泡
5’ 3’
3’
3’
5’
5’
RNA-pol (核心酶)-DNA -RNA
(四)转录终止 原核生物存在两类终止方式
1.非依赖Rho (ρ)的转录终止子
序列特征:回文序列富含GC,且在下游为连续的A
2.依赖Rho的转录终止子
序列特征:回文序列GC含量相对较少,下游序列无明显 特征
5’
3’
3’
5’
依赖 Rho因子的转录终止
◆Rho因子是rho基因的产物,广泛存在于原核和真 核细胞中,由6个亚基组成,分子量300KD。Rho因 子结合在新生的RNA链上,由5’端向3’端利用ATP 水解供能快速移动 ◆ Rho因子有ATP酶的活性和解螺旋酶的活性
DNA ρ因子
RNA聚合酶 RNA
第三节 真核生物RNA的生物合成
反式作用因子中,直接或间接结合RNA聚 合酶的,则称为转录因子(transcriptional factors, TF)
RNApol II 的转录因子
TF II D TBP(TATA盒结合蛋白) + TAFs(TBP协同因子 )
—— 结合在DNA小沟(其他DNA结合蛋白为大沟),识 别和结合核心启动子(TATA盒和Inr)
启动子
调控序列
5 3
RNA-pol
结构基因
3 5
目录
启动子
RNA聚合酶结合模板DNA的部位,称为启 动子(promoter)
RNA聚合酶 保护法
RNA聚合酶保护区 结构基因
5
3
3
5
5
3
-50 -40 -30 -20 -10 1 10
3
5
-35 区 TTGACA AACTGT
原核生物启动子保守序列
目前还不清楚RNA聚合酶Ⅱ基因的精确终止位点 (2)AAUAAA序列:
初始转录物3’末端的保守序列
对于转录产物的准确切割及加poly(A)是必须的
起始
延伸
5’ cap 5’ cap
AAUAAA AAUAAA
RNA内切酶 AAUAAA识别因子
Poly(A)聚合酶 An
Roger D. Kornberg
TATA
-40 -30 -20 -10
+10 +20
TBP的作用----定位因子 a、在TATA框处与DNA
结合
b、是所有三种RNA聚合 酶转录起始所需的因 子
TBP的作用机制: a 、两个结构域形成一个完全二元对称的马鞍型 结构,与DNA小沟有效结合
b、TBP 与DNA 结合,使DNA 弯曲了约80, TATA 盒向大沟弯曲,拓宽了小沟。
(2) TFIIA
含有至少3个亚基 与TFIID结合,稳定TFIID-DNA复合体;可能通过解除
TAFs的抑制而激活TBP
TF II A
(3) TFIIB 覆盖靠近起始点的启动位置,C端与TFIID和DNA的复合
物结合,N-端与TFⅡF协同作用募集RNA聚合酶II
TFIIB与TFIID结合,并为RNA聚合酶结合起一个桥梁作
结合,这种构象适于转录的起始 (2)CTD磷酸化可使RNA聚合酶II与DNA的
结合变得松弛,形成适于延伸的构象
RNA聚合酶II自身不能起始转录,需要依靠转
录因子的协助
转录因子
能直接、间接辨认和结合转录上游区段 DNA的蛋白质,现已发现数百种,统称为反 式作用因子(trans-acting factors)
目录
RNA聚合酶Ⅱ 与转录因子 羧基末端结构域(CTD): 1、位置:RNA聚合酶Ⅱ最大亚基羧基末端
2、结构特点: (1)具有7个氨基酸(Tyr-Ser-Pro-Thr-Ser -Pro-Ser)
的重复序列(酵母:重复26次;哺乳类:52次 )
(2)多个磷酸化位点:Ser、Thr
CTD磷酸化对调控基因转录有重要作用: (1)CTD去磷酸化,RNA聚合酶II易与DNA
4. 亚基脱落
(二)转录延长
1. 亚基脱落,核心酶变构,与模板结合松弛 ,沿着DNA模板前移
2. 在核心酶作用下,NTP不断聚合,RNA链 不断延长
(NMP) n + NTP (NMP) n+1 + PPi
转录空泡(transcription bubble):也称转录复合物,是由 RNA聚合酶的核心酶、DNA模板和转录产物RNA三者结合在 一起的复合体
目录
5′···GCAGTACATGTC ···3′ 编码链 3′··· c g t g a t g t a c a g ···5′ 模板链
转录 5′···GCAGUACAUGUC ···3′
翻译
mRNA
N······Ala ·Val ·His ·Val ······C蛋白质
目录
结构基因
5
编码链
3
模板链
转录方向
转录方向
模板链
3
编码链
5
不对称转录
(asymmetric transcription)
• 在DNA分子双链上某一特定区段,一股链 用作模板指引转录,另一股链不转录
• 模板链不总是在同一条单链上
二、RNA聚合酶
1、合成机制与DNA-pol相似 2、不需要引物
RNA聚合酶
(二)原核生物的RNA聚合酶
5 m7GpppGp…
帽子的作用:
增强mRNA的稳定性,防止被核酸酶水 解
蛋白质合成时:促进核糖体与mRNA的 结合,启动蛋白质的合成
(二)3’-末端:polyA(尾)
尾巴的作用: 参与mRNA从核内向胞质的定向转移 防止外切酶的水解,增强mRNA的稳定性
(三)mRNA的剪接 hnRNA
• 核内的初级转录产物。称为杂化核RNA (hetero-nuclear RNA, hnRNA)或核内不均一 RNA
TF II E —— 扩大DNA覆盖区至+30
TF II H 和TF II J —— H有激酶活性, 使Pol II 的CTD 磷酸化 ,PolⅡ 离开启动子区
RNA PolⅡ基因转录的过程 1、转录起始
(1) TFIID:
TBP(TATA盒结合蛋白)
因子 )
+ TAFs(TBP协同
TBP TAFs
回文序列
终止子: DNA模板上能够在转录即将结束时,提供终止信号的序 列,称为终止子(terminator)。通常为一段被间隔开的回文序列 ,存在于RNA pol已经转录过的序列中
RNA-pol
5
3
3
5
5´pppG
茎环结构使转录终止的机理 • 使RNA聚合酶变构,阻碍前移;
• 使杂交双链易于解离,转录复合物解体,RNA 产物释放
TF II A —— 含数个亚基,可能通过解除TAFs的抑制而激活TBP
TF II B —— 覆盖靠近起始点的启动位置,C端与TFIID和DNA 的复合物结合,N-端与TFⅡF协同作用募集RNA聚 合酶II
TF II F ——结合Pol II并带向启动子;RAP74(ATP依赖性解 旋酶),RAP30(与细菌因子有同源性)