酵母菌遗传课件
酵母菌遗传课件
(2)ARS-结合蛋白
酿酒酵母复制的起始可能是通 过蛋白质与ARS相互作用而引 起的 。 与 ARS结合的蛋白有六种,分 子质量分别:120kD、72kD、 62kD 、 57kD 、 53kD 和 50KD, 这些蛋白以复合物的形式存在 并与 ACS结合,这种蛋白质复 合体称为起始识别复合物 (ORC)。 ORC 与 A 区结合,也与 B 区结 合。另外,Abflp也是一种主要 的 ARS结合的蛋白,能提高复 制效率。
一、酵母菌的基因组和染色体
二、酵母线粒体基因组及其遗传
三、酵母菌中的质粒
四、酵母基因表达的调控
五、接合型基因及其基因型转换
六、酵母菌的载体系统 七、酵母双杂交系统
第一节 酵母菌的基因组和染色体
一、酵母菌的基因组
1. 酿酒酵母的单倍体细胞含有16条染色体,总长度为12068kb,其中第Ⅰ条染色 体最短(230kb ),第Ⅳ条染色体最长(1532kb) 。
(3) ARS启动染色体复制的活性 酿酒酵母基因组中约有400个ARS,但并不是所有的ARS在原染色体上都具有 自主复制活性。可是,将这些染色体上的ARS克隆到质粒上后,却都有自主复 制活性。 S 前期
从使用的时序来看,可分为两类:
S 后期
位于不同位点表现的活性不同,靠近端粒的ARS,一般不表现活性。
(1) 自主复制序列ARS的结构 • 酿酒酵母中,ARS是长度为100~200bp、富含 A T 的DNA片段。 • 根据其在质粒中的稳定性,可分为三个结构域:其中A和B最为重 要
A结构域: 保守序列,称为ARS共有序列(ACS)。 所有ARS都含有一个完全相同或非常相似的ACS。ACS内单一碱基 的突变能降低或消除其起始功能。 B结构域: 位于ACS的3’末端,长度约80bp。 C结构域: ACS 的5‘末端,富含 AT ,C结构域间不具有同源性,也不 含共有序列。
第一篇酵母菌ppt课件

-
51
作业
1、简述芽殖的过程。 2、简述子囊孢子形成的过程。 3、如何区分酵母菌的菌落与细菌的
菌落。
-
52
酵母菌的特点
①个体多以单细胞状态存在; ②多数出芽生殖,也有裂殖; ③能发酵糖类产能; ④细胞壁常含甘露聚糖; ⑤喜在含糖量较高的偏酸性环境中生长。
-
53
后面内容直接删除就行 资料可以编辑修改使用 资料可以编辑修改使用
-
55
多边芽殖,细胞 圆形、椭圆形、 卵形和腊肠形。 有假菌丝,有的 有真菌丝。
子囊孢子帽形、土星形、圆形、半圆形, 表面光滑。
是酒类酵母的污染菌。
-
50
4、假丝酵母属(Candida berkhout)
细胞圆形、卵形或长形。多边芽殖,形成 假菌丝,也有真菌丝,可生成厚垣孢子。
热带假丝酵母
产朊假丝酵母
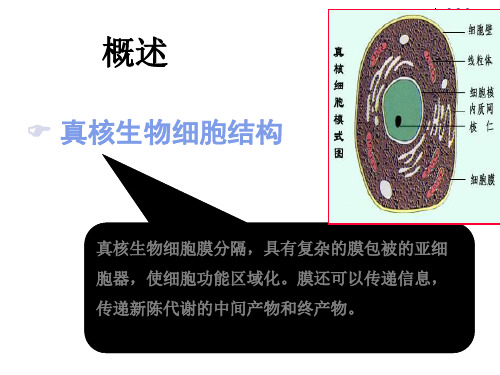
真核。 遗传信息的主要贮存库。
核仁
核膜
核基质
-
22
4、细胞器
(1)、线粒P7体1
(2)、液泡 (3)、微体 (4)、内质网
-
23
三、酵母菌的群体特征
-
24
1、固体培养基培养特征(菌落)
形状:圆形,大而厚 表面:光滑有突起或皱缩 质地:粘稠、湿润、透明或较透明、易挑取、质 地均匀 颜色: 颜色一致,多为乳白色,少数红色 边缘:整齐
在光学显微镜下,酵母菌细胞内的某些结构 模糊可见。
-
15
二、酵母菌的细胞结构
-
16
中心体 液泡 溶酶体
细胞核
中心粒
高尔基体 胞饮作用
细胞质 微丝
-
线粒体
表面突起 核糖体 微管 线粒体 胞饮作用 液泡 光滑内质网 溶酶体
酵母菌的介绍精品PPT课件

目录
• 生物定义 • 生理 • 特征 • 生殖 • 酵母菌的生长条件 • 产品种类 • 酵母菌的作用 • 危害
生物定义
酵母菌是一些单细胞真菌,并非系统演化分类的 单元。酵母菌是人类文明史中被应用得最早的微 生物。可在缺氧环境中生存。目前已知有1000 多种酵母,根据酵母菌产生孢子(子囊孢子和担 孢子)的能力,可将酵母分成三类:形成孢子的 株系属于子囊菌和担子菌。不形成孢子但主要通 过出芽生殖来繁殖的称为不完全真菌,或者叫 “假酵母”(类酵母)。目前已知大部分酵母被 分类到子囊菌门。酵母菌在自然界分布广泛,主 要生长在偏酸性的潮湿的含糖环境中,而在酿酒 中,它也十分重要。而且猫吃了还会胀大,非常 的危险。
❖ 染色体外DNA
❖
主要有两类,即线粒体DNA和2μm质粒DNA。线粒体DNA是双链DNA,
编码大量呼吸酶。
❖ 细胞质和细胞器
❖
细胞质位于细胞膜内,是一种粘稠液体,内含各种细胞器,如线粒体、
内质网、核糖体、微体、液泡。
特征
❖ 多数酵母可以分离于富含糖类的环境中,比如一些水果 (葡萄、苹果、桃等)或者植物分泌物(如仙人掌的 汁)。一些酵母在昆虫体内生活。酵母菌是单细胞真核 微生物。酵母菌细胞的形态通常有球形、卵圆形、腊肠 形、椭圆形、柠檬形或藕节形等。比细菌的单细胞个体 要大得多,一般为1~5微米或5~20微米。酵母菌无鞭毛, 不能游动。
小芽,芽细胞长到一定程度,脱离母细胞继续生长,而后形成新个体。有多
边出芽、两端出芽、和三边出芽。
❖ 分裂生殖
❖
少数种类的酵母菌与细菌一样,借细胞横分裂而繁殖。
❖ 芽裂
❖
母细胞总在一端出芽,并在芽基处形成隔膜,子细胞呈瓶状。这种方式
微生物遗传第九章酵母菌遗传

将特定基因插入到酵母菌基因组的特定位置,以研究 基因表达和调控。
基因定点突变技术
通过寡核苷酸引物或锌指核酸酶,对特定基因进行定 点突变,以研究蛋白质结构和功能。
酵母菌功能基因组学研究
全基因组表达谱分析
01
通过高通量测序技术,对酵母菌全基因组表达情况进行检测和
分析,以研究基因表达调控机制。
蛋白质组学研究
02
对酵母菌蛋白质表达、修饰和相互作用进行研究,以揭示蛋白
质功能和调控机制。和变化进行研究,以揭示代谢
途径和调控机制。
酵母菌与其他生物的基因交流与进化
基因转移与重组
研究酵母菌与其他生物之间基因的转移和重组,以揭示基因进化 机制。
基因共进化
研究不同生物之间基因的共进化关系,以揭示生物协同进化的规 律。
生物质转化
酵母菌可以将木质纤维素等生物质转化为燃料和化学品。通过基因工程手段改良酵母菌的木质素降解酶系和代谢 途径,可以提高生物质的转化效率。例如,利用基因工程技术提高酵母菌对木质素的降解能力,可以用于生物质 转化的工业生产。
THANKS
感谢观看
酵母菌遗传
• 酵母菌概述 • 酵母菌的遗传基础 • 酵母菌的基因操作 • 酵母菌遗传学研究进展 • 酵母菌遗传学应用前景
01
酵母菌概述
酵母菌的形态与分类
形态
酵母菌通常为单细胞,呈圆形、椭圆 形或圆柱形,直径一般为2-3微米。
分类
酵母菌属于真菌界,是单细胞真菌, 有数百种之多,主要分为酿酒酵母、 毕赤酵母、假丝酵母等。
要点三
药物筛选
酵母菌在药物筛选中也具有应用价值 。通过基因工程技术构建能够模拟人 类疾病的酵母菌模型,可以用于新药 筛选和药物作用机制研究。例如,利 用酵母菌模拟帕金森病等疾病模型, 用于药物筛选和机制研究。
酵母菌PPT课件
• 既处理了废水又可得到单细胞蛋白,假丝酵母可 利用味精废水生产 SCP ,蛋白质含量达 60 %,可 作为动物饲料。还有的可利用造纸厂的废水生成 SCP。
酵母菌也常给人类带来危害。腐生型酵母菌 能使食物、纺织品和其他原料腐败变质,少数嗜
高渗压酵母菌可使蜂蜜、果酱败坏;有的是发酵
工业的污染菌,影响产品质量。某些酵母菌可引 起人和植物的病害,可引起皮肤、粘膜、呼吸道、 消化道等多种疾病。
细菌一样,借细胞横分裂而繁殖。
无性孢子
节孢子( arthrospore ):
掷孢子(Ballistospore):孢子弹射
厚垣孢子(Chlamydospore):有的酵母菌在假菌 丝顶端产生的具有厚壁的孢子
掷孢子:在卵圆形营养细胞上生出小梗; 梗上形成孢子;成熟后通过特有喷射机制 射出。
节孢子:地霉属在培养初期菌体为完整的 多细胞丝状,在培养后期从菌丝内横隔处 断裂,形成短柱状或筒状,或两端钝圆的 细胞,称为节孢子。
液泡
1)单层膜包裹的细胞器;含有 机酸、盐类和水解酶类。含有 储藏物质(糖原、多磷酸盐等) 2)调节渗透压; 与细胞质进行物质交换; 3)为细胞成熟的标志
酵母菌菌落形态
与细菌菌落相似,表面湿润,粘稠, 易挑取,大而厚,颜色多为白色,少 数为红色,若培养时间太长,其表面 可产生皱褶。一般有酒香味。
3.酵母菌与人类的关系
酒类生产; 馒头面包制作; 乙醇甘油发酵; 石油及油品脱腊; 饲用药用食用SCP; 维生素、有机酸; 活性物质提取; 微生物学研究; 真核表达系统; 人类疾病;
发酵型 食用:酿酒、发面 药用:生产维生素
发酵——厌氧发酵与好氧氧化
酵母菌的遗传变异及其相关基因

酵母菌的遗传变异及其相关基因酵母菌是一种单细胞真菌,广泛存在于自然界中,包括土壤、水体、植物表面以及消化系统中等。
酵母菌在生物学研究中具有重要作用,是模式生物之一,几乎所有的有关基因表达和生化代谢调控的实验方法都可以在酵母菌上开展。
而酵母菌的遗传变异是构建酵母菌基因网络的必要过程,进而推动细胞生长和代谢的实现。
1. 酵母菌的遗传变异在自然界和实验室中,酵母菌可通过遗传变异来适应不同环境。
遗传变异包括基础性的点突变和复杂的染色体水平变异等。
其中,点突变是最常见的遗传变异形式,也是人们对遗传变异研究的重点。
点突变可分为错义突变和无义突变两类:错义突变是因为DNA序列发生了变化,让该基因表达的蛋白质发生了氨基酸的变异,从而使蛋白质结构和功能发生了改变;而无义突变则是该DNA序列变异后终止密码子生成一条截断的蛋白质链,导致细胞失能或者细胞死亡。
除此之外,酵母菌还可通过发生基因重组的方式来形成染色体水平的遗传变异。
这种变异形式通常会改变基因组的规模和染色体结构,并产生新的基因型或表型。
研究者们已通过染色体水平变异实现了酵母菌的进化人工控制,例如弱化或强化单一环境下的竞争能力、对环境毒性的适应能力等。
2. 酵母菌遗传变异相关基因(1)交叉交叉是酵母菌中基因重组的一种形式。
当酵母菌进行有性繁殖时,交叉会对基因组进行一定的乱序,从而生成不同的基因型。
正常的交叉作用需要受到遗传信号的诱导,这些信号来自酵母菌营养环境、细胞周期等不同因素。
恰当地配置这些因素可以控制交叉率,让繁殖酵母菌的下一代具有更加多样化的基因组。
事实上,随着研究进程不断深入,越来越多的基因被证明是直接或间接影响酵母菌交叉率的。
例如,环境因子下调的酵母菌细胞容易变得非常敏感,由此会减小交叉率和对外因的适应性;反之,高温和高氨基酸浓度等条件会刺激交叉率的上升。
(2)加工酵母菌的遗传变异在很大程度上是通过基因加工过程实现的。
基因加工是指对DNA进行重组、修饰、转录和翻译等一系列调节和加速基因转录的过程。
酵母菌的介绍ppt课件

种类
面包酵母
啤酒酵母
药用酵母
制造方法和性质与食品酵母相同。由于它含有丰富的蛋白质、维生素和酶等 生理活性物质,医药上将其制成酵母片如食母生片,用于治疗因不合理的饮 食引起的消化不良症。体质衰弱的人服用后能起到一定程度的调整新陈代谢 机能的作用。在酵母培养过程中,如添加一些特殊的元素制成含硒、铬等微 量元素的酵母,对一些疾病具有一定的疗效。如含硒酵母用于治疗克山病和 大骨节病,并有一定防止细胞衰老的作用;含铬酵母可用于治疗糖尿病等。
生殖
出芽繁殖 这是酵母菌进行无性繁殖的主要方式。成熟的酵母菌细胞,先长出一个 小芽,芽细胞长到一定程度,脱离母细胞继续生长,而后形成新个体。有多 边出芽、两端出芽、和三边出芽。 分裂生殖 少数种类的酵母菌与细菌一样,借细胞横分裂而繁殖。 芽裂 母细胞总在一端出芽,并在芽基处形成隔膜,子细胞呈瓶状。这种方式 很少。 有性繁殖:在合适的条件下接合子经减数分裂,双倍体核分裂为4~8个单 倍体核,形成子囊孢子,包含在由酵母菌细胞壁演变来的子囊中。子囊孢子 又可萌发成单倍体营养细胞。 酵母可以通过出芽进行无性生殖,也可以通过形成子囊孢子进行有性生 殖。无性生殖即在环境条件适合时,从母细胞上长出一个芽,逐渐长到成熟 大小后与母体分离。 有性生殖 在营养状况不好时,一些可进行有性生殖的酵母会形成孢子(一般是四 个),在条件适合时再萌发。一些酵母,如假丝酵母(或称念珠菌,Candida) 不能进行有性繁殖
繁殖大概模式
酵母菌的生长条件
营养 酵母菌同其它活的有机体一样需要相似的营养物质,象 细菌一样它有一套胞内和胞外酶系统,用以将大分子物质分解成 细胞新陈代谢易利用的小分子物质。属于异养。 水分 像细菌一样,酵母菌必须有水才能存活,但酵母需要的 水分比细菌少,某些酵母能在水分极少的环境中生长,如蜂蜜和 果酱,这表明它们对渗透压有相当高的耐受性。 酸度 酵母菌能在pH值为3.0-7.5 的范围内生长,最适pH 值为 pH4.5-5.0。 温度 在低于水的冰点或者高于47℃的温度下, 酵母细胞一 般不能生长,最适生长温度一般在20℃~30℃。 氧气 酵母菌在有氧和无氧的环境中都能生长,即酵母菌是兼 性厌氧菌,在有氧的情况下,它把糖分解成二氧化碳和水,在有 氧存在时,酵母菌生长较快。在缺氧的情况下,酵母菌把糖分解 成酒精和二氧化碳。用途 最常提到的酵母酿酒酵母(也称面包 酵母)(Saccharomyces cerevisiae),自从几千年前人类就用其 发酵面包和酒类,在酦酵面包和馒头的过程中面团中会放出二氧 化碳。 因酵母属于简单的单细胞真核生物,易于培养,且生长迅速, 被广泛用于现代生物学研究中。如酿酒酵母作为重要的模式生物, 也是遗传学和分子生物学的重要研究材料。
酵母菌ppt(上课课件)

课件教育
8
2 . 细胞膜
酵母菌的细胞膜与原核生物的基本相同。但 有的酵母菌如酿酒酵母中含有固醇类,这 在原核生物是罕见的。
课件教育
9
2 . 细胞膜的功能
▪ 调节渗透压; ▪ 选择性吸收营养物质; ▪ 分泌代谢废物。
课件教育
10
3 细胞质与细胞器
▪ 酵母菌细胞质是一种粘稠的胶体,幼龄酵 母菌细胞质稠密均匀,着色好;老龄细胞 出现液泡、空泡和储藏物质,着色不均匀。
课件教育
5
(二)酵母菌细胞结构
酵母菌的细胞 结构与其他真核生物基本相 同,包括细胞壁、细胞膜、细胞质、细胞 核及各种细胞器。
课件教育
6
1 . 酵母菌细胞壁
壁外成分: 有些菌壁外含有由多糖构成的类似荚
膜的结构。包括异多糖、甘露聚糖和淀粉 类物质。
课件教育
7
1 . 酵母菌细胞壁
▪ 细胞壁的功能: ▪ 半透性; ▪ 保护性; ▪ 与细胞合成有关。
▪ 单细胞状态存在; ▪ 出芽繁殖,也可进行裂殖或产子囊孢子; ▪ 能发酵糖类产能; ▪ 喜在含糖量高、酸度大的水生环境生长。
课件教育
4
(一) 酵母菌形态大小
▪ 酵母菌为单细胞真核微生物。 ▪ 1. 形态:因种而异,多呈圆
形,卵圆形。也有特殊形态, 如柠檬形、三角形、藕节状、 假菌丝状等 。
▪ 2. 大小:宽约1~5 mm, 长 约5~30mm 。发酵培养中 的细胞平均直径 4~5 mm.
17
三、酵母菌的繁殖方式和生活史
课件教育
18
(一)繁殖方式 无性繁殖(为主); 有性繁殖。
▪ 假酵母:只有无性繁殖过程。
▪ 真酵母:既有无性繁殖,又有有性繁殖过 程。
第九章酵母菌遗传.ppt

所有真核生物的着丝粒功能都相同,但其DNA却具有种的 特异性。
在酿酒酵母中,所有染色体的着丝粒序列(centromeric sequence,CEN)的长度均大约为130bp,由5′→3′依次分 为 CDEⅠ、CDEⅡ和CDEⅢ三个区。CDEⅠ和CDEⅢ是两个 共有序列,位于两侧,中间是CDEⅡ由 78~86个核苷酸组成, CDEⅡ的核苷酸序列中A+T含量超过90% 。
3)富含G的端粒DNA链以5’至3’延伸到染色体末端时,超出 与之互补的富含C链的12-16个核苷酸,形成一条富含G的单链, 回折形成自身G-G配对,因此它们不被核酸酶所识别,增加其 稳定性。
1987年E.Henderson等用非变性聚丙稀酰胺凝胶电泳, 吸热变性分析以及核磁共振光谱分析了端粒的顶端结构,证实 了端粒顶端回折形成发夹结构,这一结构并不是由通常的 Watson-crick配对,而是通过G-G配对而形成,富含G的特点 对形成特殊的G-G配对及染色体的复制有着重要意义。
生物 四膜虫 尖毛虫 鞭毛虫 酵母
拟南芥菜 泥毛霉菌
人
几种生物端粒的重复单位
物种
DNA来源 重复单位(5'-3')
Tetrahymena,Paramecium
大核
Stylonchia Oxytricha,Euplotes 大核
Trypanosoma,Leishmania
微染色体
Saccharomyces
二、酵母菌的染色体结构
同其它真核细胞一样,酵母菌的DNA也是与4 种主要的组蛋白结合形成核小体结构,并进一步形 成染色质。但不同的是,在酿酒酵母中没有H1组 蛋白,该蛋白在高等生物的有丝分裂过程中使染色 质维持高度超螺旋的致体的生物 学功能都严格依赖于三 种DNA序列结构:着丝 粒、端粒和复制起点, 它们是染色体功能实现 的三个基本要素。下面 就从这三方面介绍酵母 菌的染色体结构特征。
酵母菌PPT课件
2、有性繁殖
形成子囊(含有孢子的细胞)和子囊孢子。
过程:两个邻近的细胞各伸出一根管状原生质 突起,相互接触融合而成一个通道,质配,两 个核在通道内核配,形成双倍体细胞,随即减 数分裂,形成4个或8个子核,每一子核和其周 围的原生质形成孢子,称为子囊孢子。
(3)双倍体型 代表:路德氏酵母(Saccharomycodes ludwigii) 特点: 营养体为双 倍体,以出芽方 式繁殖; 单倍体子囊 孢子在子囊内发 生接合。 单倍体阶段 仅以子囊孢子形 式存在,不能独 立生活,阶段很 短。
(四)食品中常见的酵母菌
1.酵母菌属(子囊菌亚门)
2.裂殖酵母属(子囊菌亚门)
酵母菌
概述
酵母菌的形态构造
酵母菌的繁殖方式与生活史
食品中常见的酵母菌
(一)概述
泛指能发酵糖类的各种单细胞真菌, 属于子囊菌纲和半知菌纲。
1.酵母菌的特点:
单细胞
多数出芽繁殖
发酵糖类产能 胞壁含甘露聚糖 含糖量高、酸度较大的水生环境
2.酵母菌的分布
分布:偏酸的含糖环境,水果/蔬菜/蜜饯表 面和果园土壤、油田和炼油厂附近的土层(烃 类物质利用) 数量:500余种(1982) “第一种家养微生物”
(3)长与宽之比大于2。耐高渗透压,用甘蔗 糖蜜作原料时可供酒精发酵。
葡萄汁酵母(S.uvarum):它与啤酒酵母的 主要区别是全发酵棉子糖。啤酒酿造底层发酵, 饲料和药用。
食 品 微 生 物 学 刘 书 亮
Saccharomyces cerevisiae
裂殖酵母属(Schizosaccharomyces)
酵母菌遗传课件
1.着丝粒
• 概念 着丝粒是真核细胞染色体DNA上的一段特殊序列。在有丝分 裂和减数分裂时,纺锤丝与着丝粒结合,将染色体拉向细胞的两极。 • 主要类型:点着丝粒(着丝粒序列很短 酿酒酵母) 区域着丝粒(着丝粒序列较长 脉胞菌、果蝇、人 )
1.着丝粒
在酿酒酵母中,所有染色体的CEN序列的长度均大于130bp,
酵 母 菌 遗 传
酵母菌概述
酵母菌:一种单细胞真菌,属于兼性厌氧菌;个体形态有球状、卵圆、椭圆、 柱状和香肠状等。 酵母菌具有典型的真核细胞结构,有细胞壁、细胞膜、细胞核、细胞质、液泡、 线粒体等,酵母菌虽属于真核微生物,同时又具有原核生物的某些特征,例如 形成菌落特征与细菌相似,生长速度快等。 1996年完成酿酒酵母全基因组的测序,是当时完成测序的最大基因组,也是 真核生物中第一个被测序的生物。
2.GAL4转录因子和gal基因的表达调控
• 酵母半乳糖代谢酶基因(gal)表达可以被生长条件所控制。 半乳糖通过转化为6-磷酸葡萄糖后进入糖酵解途径而被酵母利用 。 • gal基因的转录是被严格调控的。gal1基因、gal7基因、gal10 基因和gal12基因在没有半乳糖的条件下不表达,而在半乳糖存 在条件下表达水平提高约1000 倍。 3.GCN4转录调控因子 GCN4能特异结合到许多氨基酸合成酶基因的启动子
1. 通用转录因子 结合在TATA序列附近,包括TFII-A、TFII-B、TGII-D、TFII-E、 TFII-F等。 2. 转录调控因子(结合在启动子上游) 转录调控蛋白由两个独立的结构域组成: (1)DNA结合区 (2)转录激活区
1.TATA 区结合蛋白
• 转录起始复合物由 TATA 元件附近序列及转录起始位点等顺式 作用元件和多种通用转录因子及RNA聚合酶Ⅱ组成。 • 成环假说
酵母菌介绍课件

氮源:氨基酸、蛋白质、尿素等
矿物质:钾、钙、镁、磷等
氧气:有氧呼吸,需要充足的氧气供应
生长因子:维生素、激素等
水分:适宜的湿度和温度
繁殖方式
酵母菌可以通过无性繁殖和有性繁殖两种方式繁殖。
有性繁殖包括接合和减数分裂,其中接合是酵母菌的主要有性繁殖方式。
无性繁殖包括芽殖和裂殖,其中芽殖是酵母菌最常见的繁殖方式。
酵母菌介绍课件
演讲人
01.
02.
03.
04.
目录
酵母菌的基本概念
酵母菌的生理特性
酵母菌在食品工业中的应用
酵母菌在生物技术中的应用
酵母菌的基本概念
酵母菌的定义
01
酵母菌是一种单细胞真菌,属于真核生物
03
酵母菌具有发酵能力,能够将糖类转化为酒精和二氧化碳
02
酵母菌广泛存在于自然界中,如土壤、空气和水中
酵母菌的繁殖方式与其生活环境和营养条件有关,不同的繁殖方式可以适应不同的环境。
酵母菌在食品工业中的应用
面包、馒头等发酵食品
酵母菌在面包、馒头等发酵食品中的作用:使面团膨胀、松软
酵母菌在面包、馒头等发酵食品中的发酵过程:酵母菌分解糖分产生二氧化碳和酒精
酵母菌在面包、馒头等发酵食品中的营养价值:富含蛋白质、维生素和矿物质
04
酵母菌在食品工业、生物技术和医学研究中具有广泛的应用
酵母菌的种类
5%
55%
30%
10%
酿酒酵母:用于酿酒和面包制作
啤酒酵母:用于啤酒酿造
面包酵母:用于面包制作
乳酸菌:用于酸奶和泡菜的制作
酵母菌的应用
面包、馒头等面点的发酵
01
酿酒工业中的发酵Байду номын сангаас
真菌遗传PPT课件

ARS 结构:
• 酿酒酵母ARS长度为100~200bp,富含AT碱基。 分为A、B、C三个结构域,A、B 2个结构域最 重要。
• 结构域A由11个核苷酸组成了保守序列,称为 ARS共有序列,简称ACS。所有的ARS中都含有 完全相同或非常相近的ACS。 ACS内单一碱基 突变就能降低或消除复制起始功能,证明它是 ARS的必须因子。但ARS的活性需要B结构域。
第13页/共29页
一、 酵母菌接合型遗传
1. 酿酒酵母生活史 • 酵母的单倍体细胞有A(a)型和α型之分。接合型分别由 matA 和 matα2个基因控制。
相同接合型酵母细胞不能发生接合。 • 酵母细胞可定期的相互转换它们的接合型,证明所有酵母细胞中同时含有matA 和
matα2个基因,但只有一类基因获得表达。
不具有体外侵染的特性,故既称为嗜杀质粒又称为类病毒颗粒。 • 外壳蛋白:M型和L型相同,由L-dsRNA编码。
第11页/共29页
• L型:4.5kb的基因组,称为L-dsRNA,含有2个 ORF, 分别编码外壳蛋白和RNA聚合酶。
• M型:含有2个相同的1.8kb RNA基因组,称为 M-dsRNA,编码毒素蛋白
见图7-13
第14页/共29页
第15页/共29页
2. 酵母接合型基因的结构
• 接合型基因MAT位于3号染色体上,两侧有两个沉默基因 HMLα和 HMRa。参与接 合型转换的基因还有HO (homothalism)。
• 这种基因转换是由HO内切酶引发的。
第16页/共29页
第17页/共29页
• 3. 酵母接合型A和α转换的盒式机制 第18页/共29页
CDE I
CDE II
CDE III
微生物遗传-8-酵母菌遗传

②
端粒
端粒(Telomere)位于 真核生物线形染色体两 端,由DNA重复序列和 蛋白质构成的复合体, 作用是保持染色体的完 整性。 端粒通常是由富含鸟嘌 呤核苷酸(G)的短的串 联重复序列组成。 端粒的重复序列不是在 染色体DNA复制时连续 合成的,而是由端粒酶 (telomerase)合成后添 加到染色体的末端。 酿酒酵母的端粒序列约 为300bp.
酵母的遗传重组即双链断裂的相对发生率 与染色体的GC丰富区相耦合,而且不同染 色体的重组频率有所差别,较小的Ⅰ、Ⅲ、 Ⅳ和Ⅸ号染色体的重组频率比整个基因组 的平均重组频率高。
② 酵母基因组遗传丰余的主要结构
DNA重复序列:如开放阅读框或者基因的间隔区包 含大量的三核苷酸重复 染色体末端重复:具有长度超过几十个kb的高度 同源区,它们是遗传丰余的主要区域 ,重组频繁。
① 镶嵌结构
多数酵母染色体由不同程度的、大范围的 GC丰富DNA序列和GC缺乏DNA序列镶嵌组成。 这种GC含量的变化与染色体的结构、基因 的密度以及重组频率有关。
GC含量高的区域一般位于染色体臂的中部, 这些区域的基因密度较高;GC含量低的区 域一般靠近端粒和着丝粒,这些区域内基 因数目较为贫乏。
Graphical View of Protein Coding Genes
Oct 11, 2008
Dec 11, 2010
Oct 31, 2011
Verified ORFs: 4726 → 4887 → 4952 Unerified ORFs: 1069 → 910 → 846 Dubious ORFs: 812 → 810
二个结构域不但可在其连接区适当部位打开, 仍具有各自的功能,而且不同两结构域可重建 发挥转录激活作用。
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
从使用的时序来看,可分为两类:
S 前期 S 后期
位于不同位点表现的活性不同,靠近端粒的ARS,一般不表现活性。
2020/4/1
---
第二节 酵母线粒体基因组及其遗传
• 线粒体是真核细胞重要的代谢中心之一。线粒体DNA(mtDNA)编 码线粒体自身所需的rRNA、tRNA以及某些蛋白质如细胞色素和 ATP酶等。
ORC与A区结合,也与B区结
合。另外,Abflp也是一种主要
的ARS结合的蛋白,能提高复
制效率。
2020/4/1
---
(3) ARS启动染色体复制的活性
酿酒酵母基因组中约有400个ARS,但并不是所有的ARS在原染色体上都具有 自主复制活性。可是,将这些染色体上的ARS克隆到质粒上后,却都有自主复 制活性。
的多肽,但是并非自给自
足。2020/4/1
---
第三节 酵母菌中的质粒(一)μm质粒 环状且周长为2μm的DNA双链、50-100个拷贝,只携带与复制和重组有关的
4个蛋白质基因(REP1、REP2、REP3和FLP),不赋予宿主细胞遗传表型, 属于隐蔽质粒。
2020/4/1
---
1.特征:质粒上有两个600bp长的反向重复序列(IR),这两个IR中间由 一个大单一区域和小单一区域所间隔。 2.在两个IR上各有一个FRT(专一性重组位点),由于这两个FRT间的相互 重组,产生A型和B型两种互变异构型的混合质粒。
基因产物或功能
a-因子 α-因子受体 分泌a-因子所需基因
α-因子 a-因子受体 α-因子加---工
第五节 接合型基因及其基因型转换
二、酿酒酵母细胞分裂的遗传控制
2、接合信息素的传递
α细胞分泌的α-因子与a细胞中 ste2基因编码的受体蛋白结合。 同样,a细胞分泌的a-因子与α细 胞中ste3基因编码的受体蛋白结 合。
4.基因组含有52个完整的Ty因子(反转座子)。
2020/4/1
---
第一节 酵母菌的基因组和染色体
1.核小体:DNA和组蛋白(H2A、H2B、H3和H4) 酿酒酵母中无H1(有丝分裂中维持染色质的高度超螺旋) 2.着丝粒(centromere):点着丝粒和区域着丝粒 3.端粒(telomere):末端重复序列和蛋白质
1996年完成酿酒酵母全基因组的测序,是当时完成测序的最大基因组,也是 真核生物中第一个被测序的生物。
2020/4/1
---
一、酵母菌的基因组和染色体 二、酵母线粒体基因组及其遗传 三、酵母菌中的质粒 四、酵母基因表达的调控 五、接合型基因及其基因型转换 六、酵母菌的载体系统 七、酵母双杂交系统
4.HAP1 HAP2和HAP3
• HAP1 HAP2和HAP3都是转录因子,控制酵母细胞色素c基 因CYC1和CYC7的转录。
• CYC1 UAS 由 UAS1 和UAS2 两个位点组成。 UAS1 与葡萄糖抑制时基因的转录有关 UAS1和UAS2共同作用 是乳糖去阻遏时基因转录所必需的。
5.α1、α2和a1调节蛋白
自1979年首次发现酿酒酵母的ARS以来,已在酿酒酵母中约有400个ARS, ARS/40kb。但这些ARS的使用频率不同,变动在0-100%之间。
2020/4/1
---
(1) 自主复制序列ARS的结构 • 酿酒酵母中,ARS是长度为100~200bp、富含 A T 的DNA片段。 • 根据其在质粒中的稳定性,可分为三个结构域:其中A和B最为重
2020/4/1
---
第五节 接合型基因及其基因型转换
二、酿酒酵母细胞分裂的遗传控制
2、接合信息素的传递
2020/4/1
---
第五节 接合型基因及其基因型转换
2020/4/1
---
第一节 酵母菌的基因组和染色体
一、酵母菌的基因组
1. 酿酒酵母的单倍体细胞含有16条染色体,总长度为12068kb,其中第Ⅰ条染色 体最短(230kb ),第Ⅳ条染色体最长(1532kb) 。
2. 基因组中没有明显的操纵子结构,有间隔区和内含子。
3. 酿酒酵母染色体基因组中,有5885个可能是编码蛋白质的ORF,每个ORF约为 1.4kb,而基因间的平均间隔为600bp。(这说明酵母基因比其它高等真核生物基 因排列紧密。)
2020/4/1
---
二、酵母转录调控因子(反式作用蛋白)
1. 通用转录因子 结合在TATA序列附近,包括TFII-A、TFII-B、TGII-D、TFII-E、 TFII-F等。
2. 转录调控因子(结合在启动子上游) 转录调控蛋白由两个独立的结构域组成: (1)DNA结合区 (2)转录激活区
2020/4/1
酵 母 菌 遗 传
-
---
酵母菌概述
酵母菌:一种单细胞真菌,属于兼性厌氧菌;个体形态有球状、卵圆、椭圆、 柱状和香肠状等。
酵母菌具有典型的真核细胞结构,有细胞壁、细胞膜、细胞核、细胞质、液泡、 线粒体等,酵母菌虽属于真核微生物,同时又具有原核生物的某些特征,例如 形成菌落特征与细菌相似,生长速度快等。
2020/4/1
---
第五节 接合型基因及其基因型转换
一、酿酒酵母的生活史
2020/4/1
---
第五节 接合型基因及其基因型转换
一、酿酒酵母的生活史 二、酿酒酵母细胞分裂的遗传控制
1、a和α单倍体细胞 2、接合信息素信号的传递 3、接合型基因MAT在接合过程中的调控作用 三、接合型基因的转换 1、酵母接合型基因的结构 2、接合型转换机理
•
抑制性小菌落:丢失了部分线粒体DNA
2020/4/1
---
(二)酵母线粒体基因组的物理图谱及其特点
1.酵母线粒体DNA是周长
约26μm的环状DNA分子,
其大小为84kb。
2.绝大多数的mtDNA中没
有重复核苷酸序列。
3.线粒体是半自主性的,
能复制并传递给后代,还
能转录所编码的遗传信息,
合成线粒体某些自身特有
要
A结构域: 保守序列,称为ARS共有序列(ACS)。 所有ARS都含有一个完全相同或非常相似的ACS。ACS内单一碱基
的突变能降低或消除其起始功能。
B结构域: 位于ACS的3’末端,长度约80bp。
C结构域: ACS 的5‘末端,富含 AT ,C结构域间不具有同源性,也不 含共有序列。
2020/4/1
裂和减数分裂时,纺锤丝与着丝粒结合,将染色体拉向细胞的两极。
• 主要类型:点着丝粒(着丝粒序列很短 酿酒酵母) 区域着丝粒(着丝粒序列较长 脉胞菌、果蝇、人 )
2020/4/1
---
1.着丝粒
在酿酒酵母中,所有染色体的CEN序列的长度均大于130bp,
由5′→3′依次分为 CDEⅠ、CDEⅡ和CDEⅢ三个区。
酵母细胞的单倍体类型是由接合型基因(MATα和 MATa)决定的。MATα编码α1和α2两个蛋白调控因子, 它们决定α型细胞的表型。MATa编码a1蛋白,控制a 型细胞的表型。
2020/4/1
---
第五节 接合型基因及其基因型转换
一、酿酒酵母的生活史 二、酿酒酵母细胞分裂的遗传控制
1、a和α单倍体细胞 2、接合信息素信号的传递 3、接合型基因MAT在接合过程中的调控作用 三、接合型基因的转换 1、酵母接合型基因的结构 2、接合型转换机理
4.复制起点:控制DNA复制起始的一段DNA序列—ARS
ARS若克隆到质粒中,可使质粒DNA在酵母中自主复制。 1)ARS的结构
A:ACS在所有的ARS都完全相同或相似 ARS B:ACS的3’末端
C:A
2020/4/1
---
1.着丝粒
• 概念 着丝粒是真核细胞染色体DNA上的一段特殊序列。在有丝分
分类:L型类病毒、M型类病毒 L-dsRNA:编码自身和M型的蛋白外壳和RNA聚合酶 M-dsRNA:编码杀伤毒素蛋白,分泌到细胞外。
2020/4/1
---
嗜杀型酵母
含L型类病毒 和M型类病毒
L型类病毒:含有 4.5kb的双链RNA
2020/4/1
M型类病毒:含有两条 1.8kb双链RNA
---
敏感型酵母
REP3
A-type
REP1
REP2
FRT
FRT
FLP
2020/4/1
REP1
B-type
REP3
---
REP2
FRT
FRT
FLP
(二) 嗜杀现象
1963年,Bevan和Makower发现酿酒酵母中某些菌株可产生毒素 而杀死其他酵母的现象。
嗜杀株(killer):产生毒素的菌株 敏感株:对毒素敏感的菌株 中性株:既不产毒素又不敏感的菌株 该特性由两种具有自我复制能力的细胞遗传因子----双链线状 RNA(dsRNA)决定的,它们通常以蛋白质外壳包裹着的粒子状态存 在于细胞之中,不具有体外侵染的特性。也称为类病毒颗粒(viruslike particle).
酿酒酵母中2/3的端粒含有1~4个拷贝的Y'。
X和Y'作用:
非必需,对端粒的稳 定、染色体断裂后的 修复以及在减数分裂 中染色体联会方面都 起辅助作用。
2020/4/1
---
3.复制起点
酵母的复制起点是指染色体上控制DNA复制起始的一小段DNA序列,通常 称为自主复制序列(autonomously replicatory sequence,ARS)。
只含L型类病毒 不含M型类病毒
不含L型类病毒 不含M型类病毒
第四节 酵母基因表达的调控
根据其性质,真核基因调控分两类: 1、瞬间调控(可逆性控制) 2、发育调控(不可逆性控制)
2020/4/1
---
一、酵母基因的启动元件(顺式作用元件)