基因定点诱变常用方法
基因定点突变方法及其应用

基因定点突变方法及其应用
基因定点突变是指在基因组中特定的位置发生的变异事件。
这些定点
突变可以是单个碱基的替代、插入或删除,也可以是染色体片段的重排和
结构变化。
基因定点突变是遗传学和分子生物学研究中的重要技术,具有
广泛的应用前景。
在基因定点突变的研究中,常用的方法包括基于随机突变和基于定点
突变的方法。
一、基于随机突变的方法:
1.化学诱变:通过化学物质如亚硫酸乙基甲酯(EMS)或N-亚硝基-N-
乙基脲(ENU)等诱导基因组范围内的突变。
这些突变通过对群体进行筛选
和筛选,从而找到目标基因的突变。
2.辐射突变:用射线如X射线、γ射线,或放射性物质如乙烯基腈,等辐射处理生物体,以诱导其基因组上的随机突变。
基于随机突变的方法广泛应用于物种、疾病、发育和进化等研究中。
它们可以帮助揭示基因功能的重要性和与特定物种和表型相关的基因。
二、基于定点突变的方法:
1.基因敲除/敲入:通过合成受损的DNA片段或外源DNA片段,将其
导入到目标基因中,从而造成其功能异常或置换为新的基因序列。
这种方
法可用于明确或验证基因的功能,并探索特定突变的表型影响。
基因定点突变技术.

四、展望
定点突变技术可以高效地应用于DNA缺失改造、 蛋白质工程和酶等多个研究领域。它不仅加深人们对 蛋白质、酶等物质的了解,而且也为进一步研究这些 物质提供技术支持。另外,突变技术可以对某一特性 相关位点同时进行突变,这就极大地提高了获取突变 子的效率,避免了高强度的筛选工作,同时也节约了 大量的时间和资金。突变技术在蛋白质工程研究将会 继续保持迅猛势头,且在医学、农业科学、肿瘤研究、 基因表达与调控等领域,突变技术的应用也越来越广 阔。因此,我们有理由相信突变技术在未来会有光明 的应用前景。
基因定点突变技术
目录
一、定点突变简介
二、基因定点诱变 三、基因定点突变应用
四、展望
一、定点突变简介
定点突变(site-directed mutagenesis) 技术可以有目的性地在已知DNA 序列中 取代、插入或缺失一定的核苷酸片段,可以 有目的或针对性的改变DNA序列中的碱基 次序;它不仅可以用来阐明基因的调控机理, 也可以用来研究蛋白质结构与功能之间的关 系。
1、寡核苷酸盒式诱变
(Cassette mutagenesis)
利用一段人工合成的具有突变 序列的寡核苷酸片段,即寡核苷酸 盒,取代野生型基因中的相应序列。
ቤተ መጻሕፍቲ ባይዱ
2、寡核苷酸引物诱变
使用化学合成的含有突变碱基的寡 核苷酸片段作为引物,启动单链DNA 分子进行复制,随后这段寡核苷酸引 物便成为了新合成DNA子链的一个组 成部分,因此所产生出来的新链便具 有已发生突变的碱基序列
1、突变技术在蛋白质工程中的应用
突变技术在蛋白质工程中的应用 非常广泛和成功。此技术不仅是蛋白 质定向改造的强有力工具,而且也是 研究蛋白质结构和功能关系的重要方 法。
基因定点诱变技术与DNA与蛋白质互作及定点突变介绍

30
酵母单杂交的基本 原理示意图
从 拟 南 芥 cDNA 文 库 中 筛选与顺式元 件DRE结 合的 转录因子示意图。
31
三)凝胶阻滞实验 (electrophoretic mobility shift assay, EMSA)
六)体内足迹试验
• 用适量DMS处理完整的游离细胞,使染色 质中的G残基甲基化,提取DNA并加入六氢 吡啶切割DNA链。与对照的裸露DNA经甲基 化处理后形成42
43
七)染色质免疫沉淀实验 Chromatin Immunoprecipitation(ChIP) Assay
33
34
• EMSA • 还被
用于 研究 与蛋 白质 相结 合的 DNA • 序列 的特
异型。
35
四)DNaseI足迹试验 (DNase I foot printing)
• 主要步骤: ① 用32P标记DNA双链末端,并用RE切去一端; ② 加入细胞特定周期蛋白质提取物,温育; ③ 加入适量DNaseI或硫酸二甲酯-六氢吡啶,使 DNA链发生断裂。 这一反应中,DNaseI或硫酸二甲酯的用量非常关键, 要保证一条链只发生一次断裂! ④沉淀DNA(包括与DNA相结合的蛋白质); ⑤进行DNA凝胶分析。
真核生物DNA序列(非编码序列)和被 转录的结构基因距离较近,和转录调控有关。
A 启动子(启动子上游近侧序列) B 增强子 C 沉默子
29
启动子(promtor)在转录起始点上游约100-200bp以内, 每个元件长度约为7-20bp,决定RNA聚合酶Ⅱ转录起始 点和转录频率的关键元件。
第七章 定点诱变

两种活性结合产生平端或较短5’突出端的缩短 分子
利用BAL 31核酸酶制备嵌套缺失突变体
处理不同时间
Klenow DNA聚合酶 补平末端
与克隆载体连接
3.DNase Ⅰ 的消化
一种内切酶,可优先水解 ds or ss DNA 中的 嘧啶核苷酸 。
基因A P2
P1
基因B
5. 大引物PCR诱变法
大引物PCR法由Kammann M等人于1989
年提出,随后经Sarkar G和Sommer S S等人加 以改进。
该方法需2个侧翼引物和1个内部诱变引物, 经过2轮PCR获得突变的全长DNA。
第1轮PCR用诱变引物和1个侧翼引物,扩增 产物经凝胶纯化后作为大引物再与另一侧翼 引物进行第2轮PCR,产生全长的突变的DNA。
第七章f cloned DNA
突变是研究基因结构与功能的最基本的手段
经典的方法:分离自发突变体
用物理、化学诱变剂处理活体
诱变-整个生物体-目的基因上的突变极其有限
体外诱变(in vitro mutagenesis):对克隆化的 DNA进行诱变处理可得到经典诱变所能得到 的所有种类的突变
突变类型: 单个碱基的置换 简单的插入或缺失 系统的缺失、插入或成串碱基的置换
第三节 寡核苷酸介导的定点诱变 p176
基因的定点诱变(site-directed mutagenesis): 使已克隆基因或DNA片段中的任何一个特定碱 基发生取代、插入或缺失突变的过程
需一个或多个含突变碱基的诱变寡核苷酸作为 引物进行DNA复制,使寡核苷酸引物成为新合 成的DNA子链的一部分
基因定点诱变技术与DNA与蛋白质互作及定点突变介绍

33
• EMSA • 还被
用于 研究 与蛋 白质 相结 合的 DNA • 序列 的特
异型。
34
四)DNaseI足迹试验 (DNase I foot printing)
• 主要步骤: ① 用32P标记DNA双链末端,并用RE切去一端; ② 加入细胞特定周期蛋白质提取物,温育; ③ 加入适量DNaseI或硫酸二甲酯-六氢吡啶,使 DNA链发生断裂。 这一反应中,DNaseI或硫酸二甲酯的用量非常关键, 要保证一条链只发生一次断裂! ④沉淀DNA(包括与DNA相结合的蛋白质); ⑤进行DNA凝胶分析。
23
24
酵母双杂交的基本原理示意图 25
二) 酵母单杂交系统
• 酵母单杂交系统是上世纪90年代中发展起来的研 究DNA-蛋白质之间相互作用的新技术,可识别稳 定结合于DNA上的蛋白质,在酵母细胞内研究真核 生物中DNA-
14
一)酵母双杂交系统 (yeast two-hybrid system)
1. 原理:
• 真核生物的转录因子大多是由两个结构上分开、 功能上独立的结构域组成的,即 DNA结合域(BD) 和转录激活域(AD)。
• 单独地BD能与特定基因地启动区结合,但不能激 活基因的转录,而由不同转录因子的BD和AD所形 成的杂合蛋白却能行使激活转录的功能。
20
4)一旦酵母细胞中表达的“诱饵”蛋白与“猎物” 载体中表达的某个蛋白质发生相互作用, 不同转录调控因子的AD和BD结构域就会 被牵引靠拢,激活报告基因表达。
5)分离有报告基因活性的酵母细胞,得到所 需要的“猎物”载体,获得与已知蛋白相互 作用的新基因。
21
22
AD:Activation domain DNA-BD:Binding domain
基因定点突变全攻略

基因定点突变全攻略一、定点突变得目得把目得基因上面得一个碱基换成另外一个碱基.二、定点突变得原理定点突变就是指通过聚合酶链式反应(PCR)等方法向目得DNA片段(可以就是基因组,也可以就是质粒)中引入所需变化(通常就是表征有利方向得变化),包括碱基得添加、删除、点突变等。
定点突变能迅速、高效得提高DNA所表达得目得蛋白得性状及表征,就是基因研究工作中一种非常有用得手段。
体外定点突变技术就是研究蛋白质结构与功能之间得复杂关系得有力工具,也就是实验室中改造/优化基因常用得手段。
蛋白质得结构决定其功能,二者之间得关系就是蛋白质组研究得重点之一。
对某个已知基因得特定碱基进行定点改变、缺失或者插入,可以改变对应得氨基酸序列与蛋白质结构,对突变基因得表达产物进行研究有助于人类了解蛋白质结构与功能得关系,探讨蛋白质得结构/结构域。
而利用定点突变技术改造基因:比如野生型得绿色荧光蛋白(wtGFP)就是在紫外光激发下能够发出微弱得绿色荧光,经过对其发光结构域得特定氨基酸定点改造,现在得GFP能在可见光得波长范围被激发(吸收区红移),而且发光强度比原来强上百倍,甚至还出现了黄色荧光蛋白,蓝色荧光蛋白等等。
定点突变技术得潜在应用领域很广,比如研究蛋白质相互作用位点得结构、改造酶得不同活性或者动力学特性,改造启动子或者DNA作用元件,提高蛋白得抗原性或者就是稳定性、活性、研究蛋白得晶体结构,以及药物研发、基因治疗等等方面.通过设计引物,并利用PCR将模板扩增出来,然后去掉模板,剩下来得就就是我们得PCR 产物,在PCR产物上就已经把这个点变过来了,然后再转化,筛选阳性克隆,再测序确定就行了.三、引物设计原则引物设计得一般原则不再重复.突变引物设计得特殊原则:(1)通常引物长度为25~45 bp,我们建议引物长度为30~35 bp。
一般都就是以要突变得碱基为中心,加上两边得一段序列,两边长度至少为11—12 bp。
若两边引物太短了,很可能会造成突变实验失败,因为引物至少要11-12个bp才能与模板搭上,而这种突变PCR要求两边都能与引物搭上,所以两边最好各设至少12个bp,并且合成多一条反向互补得引物。
基因的定点诱变

因为DNA复制酶有校正功能及DNA复制 后修复系统,正常大肠杆菌自发突变频 率较小。 与校正和修复有关的基因突变后细胞内 基因突变频率大大增加,这样的菌株叫 突变菌株。
流程: 把携带待突变基因的质粒转入增变菌株 扩增,------随机突变体库。 把该库的重组质粒转化到正常宿主中, 筛选鉴定突变体。
寡核苷酸介导的定点诱变
①基本原理: 使用化学合成的含有突变碱基的 寡核苷酸片段作为引物,启动单链DNA 分子进行复制,随后这段寡核苷酸引物 便成为了新合成DNA子链的一个组成部 分,因此所产生出来的新链便具有已发 生突变的碱基序列
作为诱变剂的寡核苷酸序列:
人工合成一段寡核苷酸序列,该序列中 除了所需的碱基突变外,其余的则与目 的基因编码链的特定区段完全互补 错配碱基的位置应设计在寡核苷酸分子 的中央部位,每侧至少10-15个碱基 通常采用化学合成 无回文,重复和自身互补序列
即使获得期望表型的突变体,无法保证 突变确实发生在目的基因上 在基因克隆和核酸测序技术发展之前, 无法知道基因中突变的位置和性质
Directed evolution (定向进化或直接进化)
对目的基因人为制造大量突变,然后 按照特定的需要和目的,给予选择压力, 反复诱变,循环筛选,将满足要求的、适 合特定目的的分子筛选出来,获得满足需 要的性能改良的蛋白质,从而实现在试管 中分子水平的模拟进化。
基因家族的改组
交错延伸重组 图10-5
以两个以上的有一定同源性的DNA片段 为模板进行PCR反应,通过交换模板机制 实现DNA序列的重新组装。 不需要DNaseI切割DNA,简化了程序。
随机引发重组 图10-6
分子生物学--定点诱变与蛋白质工程课件

❖ N端-----------笨丙-天冬-----------谷-甘-------------- C端 多肽链
三、定点诱变的原理
定点诱变原理示意图
❖ The Nobel Prize in Chemistry 1993
"for contributions to the developments of methods within DNA-based chemistry"
2.诱变过程
1)合成含有目的 基因的正链DNA; 2)合成含有特殊 突变碱基的引物; 3)制备异源双链 DNA(右图); 4)富集和转化双 链DNA分子; 5)筛选突变体并鉴定
(二)Kunkel定点诱变法
❖ 体外DNA合成往往是不完全的,所以部分合 成的DNA分子必须通过蔗糖密度梯度离心除 去,获得纯化的突变DNA。
❖ 理论上来说,DNA是半保留复制的,应用寡 核苷酸定点诱变时,所形成的噬菌体中携带 突变基因的应为一半。但实际上,由于技术 上的原因通常只有1-5%的噬菌斑含有突变基 因的噬菌体。
❖ 因此,为了获得更多含有突变噬菌体的噬菌 斑,必须提高突变体的比率。目前已有多种 改良的寡核苷酸定点诱变方法,此处将 Kunkel 1985年建立的方法做以简单介绍。
定点诱变技术

PCR介导的基因突变
在基因5’和3’末端产生突变 重叠延伸PCR 大引物PCR法
在基因5’和3’末端产生突变
重叠延伸法定点突变技术的原理示意图
重叠延伸PCR法小结
缺点:需要2对引物,进行3次PCR,并且需要对中 间产物进行纯化。 优点:几乎没有特殊限制,而且成功率高,因此运 用非常广泛。
此法的成功关键,是要得 到好的含U单链模板DNA。
噬菌粒载体 (phagemid )
M13K07辅助感染 产生带 U 的单链 DNA
无5‘-3’外切活性 3‘-5’外切活性低
这种方法得到的突变率太低,约为1-5%。主要原因是含突变 位点的双链DNA,转入E.coli后,被其修复系统修复了。
盒 式 定 点 突 变
DNA Shuffling
外显子、单基因和基因家族的重组装 随机引物延伸法 交错延伸法
定点突变
点突变——碱基删除、增补和替换
易错PCR(epPCR)
How DNA shuffling works
How DNA shuffling is done in the tube
Random fragmentation of a pool of related genes; Self-priming polymerase reaction and template switching (causing crossovers); PCR amplification with primers of reassembled products
在正常情况下,尿嘧啶N-糖基化酶(ung+)可以去除掺 入DNA中的尿嘧啶残基。但 在ung-的菌株中,此酶失活。
在大肠杆菌dut- ung-菌株 中生长的M13噬菌体的单链基 因组DNA中将含有20-30个尿 嘧啶残基。用这些噬菌体感染 ung+菌株,尿嘧啶被迅速去 除,DNA链遭到破坏,感染 力下降约5个数量级。
基因工程第七章DNA定点诱变

• 3’端所形成的杂交体足以引导DNA合成。如果
错配核苷酸太靠近3’端,3’端将不能形成稳定的 杂交体,易被外切活性降解。所以3’端需有79bp完全配对;
• 为便于筛选,应选用可形成稳定杂交体而长
度最短的诱变寡核苷酸。一般17-19bp, 错配在 中央,使得完全配对的杂交体与错配杂交体 之间的热稳定性差异足够大。
位点选择定点诱变法
三、Transformer SiteDirected mutagenesis
转化子诱变法
Synthesize second strand
Digest DNA (primary digestion)
Transform E.coli mutS to propagate plasmids
ATG ATG
ATG ATG
BamHI
ATG
BamHI
PCR ATG
ATG
EcoRI
PCR
ATG
EcoRI
重叠延伸
BamHI
ATG
PCR EcoRI
第二节 嵌套缺失
第二节 嵌套缺失
1. 外切核酸酶III的消化
Exonuclease III 5‘ 3‘
5‘ 3‘
5‘ 3‘
2. BAL 31的消化
DNA ligase
U
U
U
U
U
2.原理
Insert target DNA
转化 E.coli CJ236
U
U
U U
U
U
Isolate phagemid ss DNA 与突变寡核苷酸退火
转化E.coli MV1190 (dut+/ung+)
UU
基因定点突变DNA实验技术方法

基因定点突变DNA实验技术方法要研究和检测基因定点突变,需要使用一系列的实验技术方法。
以下是几种常用的DNA实验技术方法:1.基因测序技术基因测序技术是研究基因组突变的关键方法之一、它可以将DNA分子按顺序解码,并确定单个核苷酸的序列。
经过多年的发展,现在有很多种基因测序技术可供选择,如Sanger测序、Illumina测序、PacBio测序和单分子DNA测序。
这些技术可以高效地测定基因组中各个位置的核苷酸序列,并揭示基因定点突变的存在。
2.聚合酶链式反应(PCR)PCR是一种用于扩增特定DNA片段的方法,可以在PCR反应过程中选择性地扩增感兴趣的基因片段。
对于基因定点突变的研究,PCR可以用来扩增包含突变位点的DNA片段,并通过测序分析来确定突变的类型。
3.引物延伸法引物延伸法是一种常用的检测单核苷酸多态性(SNP)和点突变的方法。
该方法使用引物和DNA聚合酶来识别特定位点,并从该位点延伸引物,以确定突变的存在与否。
引物延伸法可以用于快速、高效地检测多个位点的突变。
4. 限制性酶切酶(Restriction Enzyme Digestion)限制性酶切酶可以通过识别特定的DNA序列并在该序列周围切割DNA来检测和分析基因定点突变。
当突变中产生或消失限制性酶切位点时,可以通过酶切后的DNA片段的大小变化来检测突变。
5. DNA芯片技术(DNA Microarray)DNA芯片技术是一种高通量的基因分析技术,可以同时检测和比较大量的基因表达水平。
通过将DNA分子固定在芯片的表面并用荧光探针进行杂交反应,可以检测不同样品中基因定点突变的存在。
以上是几种常用的DNA实验技术方法,用于研究和检测基因定点突变。
随着技术的不断发展和创新,这些方法将进一步提高灵敏度和分辨率,为基因定点突变的研究提供更多的选择和可能性。
PCR诱变

2 易错PCR(error-prone PCR)
DNA聚合酶在进行扩增目的DNA时会以一定的 频率发生碱基错配,这一现象恰好提供了一 种特定基因进行随机诱变的可能方法。
利用PCR过程中出现的碱基错配进行特定基因 随机诱变的技术就称为易错PCR。 原理:较低的聚合酶(如Taq酶)在特定措施 下很容易向扩增产物中掺入随机突变
经PCR后,每个管产生了含有改变了特定碱基 的产物,但留有缺口。因此,产物是线状的 DNA链,由于引物互补位置不同,两个管的产 物的缺口位置也不同,当把两个反应管的产 物混合,经变性和退火处理,来自不同反应 管的线状DNA链可杂交形成双链环状的DNA分 子,但仍有缺口。由于环化分子可以转化大 肠埃希菌,在体内可把缺口修复,这样可以 获得定点改变了特低碱基的基因。
扩增战略:
三、 PCR随机诱变
当对目的序列了解很少的情况下,需 要引入大量的随机诱变,构建突变库,再 逐个筛选和鉴定 策略 1、利用简并引物 2、易错 PCR (error-prone PCR )
1、利用简并引物
简并引物的特点 5’端有十几个碱基是随机组合的。这一方 法除引物特殊外,其余均与定点突变技术完 全一样。利用简并引物可以产生大量随机突 变,并且这些突变都集中在很窄的一段区域 内。
2、大引物PCR法(megaprimer PCR )
先设置一个PCR反应产生一个含突变的DNA 片段,然后再以此DNA片段作为引物与原模板 退火进行PCR扩增得到含突变的完整的基因。 因为作为引物使用的DNA片段较通常的引物要 大许多甚至有上百碱基,所以命名为大引物 PCR法。
大引物PCR 定点诱变技术路线
ቤተ መጻሕፍቲ ባይዱ
1、重叠延伸PCR (over-lap extension PCR, OE-PCR ) 2、大引物PCR法(megaprimer PCR ) 3、环状诱变 PCR 法 4、TAMS 定点诱变技术
基因定点诱变
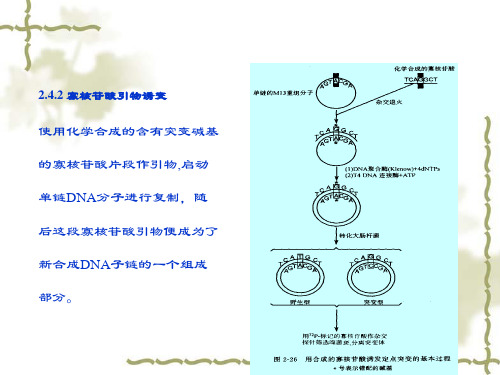
使用化学合成的含有突变碱基 的寡核苷酸片段作引物,启动 单链DNA分子进行复制,随 后这段寡核苷酸引物便成为了 新合成DNA子链的一个组成 部分。
寡核苷酸引物诱变过程
正链DNA合成 突变引物合成 异源双链DNA分子制备 闭环异源双链DNA分子富集 转化 突变体筛选
寡核苷酸引物诱变法的局限性
异源双链DNA分子并非真正异源 突变体子代中突变碱基被错配修 复体系修复
提高寡核苷酸引物诱变突变效率的方法
(1) Kunkel 定点诱变法: 1985年由
(2) 硫代磷酸诱变法: 已知有些限制酶不能切割 硫代磷酸DNA分子.在异源双链DNA分子 制备时,加入硫代核苷酸,使之掺入突变 链中.然后用前述酶切割,并用外切酶局 部消化后,进行聚合反应,从而产生具有 定点突变的异源双链DNA
2.4.3 PCR诱变
(1) 重组PCR定点诱变:克服寡 核苷酸引物诱变能力仅限于5’ 端.
特点:经3轮PCR反应,一对互补 的带有突变碱基的内侧引物和 2个外侧引物
(2) 大引物诱变:核心是第一 轮PCR产物为第二轮PCR引 物
返回目录
返回第二章
定点突变技术

操作:设计引物,分别PCR,前两次PCR 反应产物经琼脂糖凝胶电泳鉴定后无需 纯化,直接将胶条切下置于EP管中, 80℃冷冻10min,然后将胶条离心后分 别取上清液作为模板进行第三次PCR,以 获得全长的突变目的基因
PCR介导的定点突变法其优点是操作较简 单,突变的成功率可达100%。但它亦 有两个缺点:①后续工作较复杂,PCR扩 增产物通常需要连接到载体分子上,然
寡核有酸引物介导的定点突变 法其优点是保真度比PCR突变法 高,经过改进后使该方法突变少 成功率大大提高,缺点是操作过 程环节复杂制。
盒式突变法具有简单易行、突变效率高 等优点,还可以在一对限制酶切位点内 一次突变多个位点。缺点是合成多条引 物的成本较高。另外,在一般情况下, 在靶DNA片段的两侧往往难以满足存在 一对限制性酶切位点的要求,限 制了该 方法的广泛应用。然而一旦具备了这样 的条件该方法则为首选。
定点突变技术
刘微
基因的定点突变技术
点突变的技术有很多种,常见的有: ㈠寡核苷酸介导的定点突变技术
(M13噬菌体法) ㈡ 盒式诱变 ㈢PCR点突变技术
1.引物PCR定点诱变法 2.重组PCR定点诱变法 3.重叠延伸PCR技术
寡核苷酸引物诱变技术 (M13噬菌体)
噬菌体M13的生活周期有二个阶段,在噬菌体 粒子中其基因组为单链,侵入宿主细胞以后, 通过复制以双链形式存在。将待研究的基因 插入载体M13,制得单链模板,人工合成一 段寡核苷酸(其中含一个或几个非配对碱基) 作为引物,合成相应的互补链,用T4连接酶 连接成闭环双链分子。经转染大肠杆菌,双 链分子在胞内分别复制,因此就得到两种类 型的噬菌斑,含错配碱基的就为突变型。
重组PCR定点诱变2
操作:设计引物,分别PCR, 从两个PCR反 应管中各取出3μl PCR反应产物,混匀后用 CaCl2转化法转化至感受态大肠杆菌中。涂 平板后,从转化的细菌菌落中随机挑选若干,筛 选。
基因定点诱变的方法及原理

基因定点诱变的方法及原理基因定点诱变是指在特定位置引发基因突变的一种技术或方法。
通过基因定点诱变技术,可以精确地改变基因组中特定位置的碱基序列,从而研究或改变目标基因的功能。
目前常用的基因定点诱变方法主要有以下几种:1. CRISPR-Cas9系统:CRISPR-Cas9(Clustered Regularly Interspaced Short Palindromic Repeats-CRISPR associated protein 9)是一种基于RNA-DNA 相互识别的靶向基因编辑技术。
该系统利用Cas9蛋白通过结合到特定的DNA 序列来导向编辑目标位置,而CRISPR RNA(crRNA)和互补序列的转录过程产生了指导RNA(sgRNA)。
CRISPR-Cas9系统可以通过设计合成特定的sgRNA来诱导Cas9蛋白与目标基因的DNA序列结合,并在目标位点引入双链断裂,通过自然修复过程来实现基因突变。
2. TALEN系统:TALEN(Transcription activator-like effector nuclease)是一种由TAL(Transcription activator-like)蛋白和核酸酶融合而成的基因编辑工具。
TAL蛋白可通过识别和结合特定的DNA序列来实现靶向基因编辑。
TALEN 系统利用设计合成的TAL蛋白与核酸酶的融合体结合到目标基因的DNA序列上,并通过酶活性诱导DNA的双链断裂,从而引发基因突变。
3. ZFN系统:ZFN(Zinc finger nuclease)是由锌指蛋白(Zinc Finger Protein)与核酸酶(nuclease)融合而成的一种基因编辑工具。
锌指蛋白能够识别和结合到特定的DNA序列上,而核酸酶则通过识别锌指蛋白与DNA结合后的底物序列引发DNA的切割。
ZFN系统利用设计合成的锌指蛋白与核酸酶的融合体结合到目标基因的DNA序列上,从而在特定的位置诱导DNA的双链断裂,进而引发基因突变。
第七章基因的定点诱变

野 生 型 DNA
H in d I I I 移走小片段
E coR I
H in d I I I E coR I
人工合成的寡核苷酸片段 退火
H in d I I I
H in d I I I
E coR I
突变体序列
E coR I
D N A连 接 酶
简 单 盒 式 取 代 诱 变
寡核苷酸介导的定点诱变
①基本原理: 使用化学合成的含有突变碱基的
寡核苷酸片段作为引物,启动单链DNA 分子进行复制,随后这段寡核苷酸引物 便成为了新合成DNA子链的一个组成部 分,因此所产生出来的新链便具有已发 生突变的碱基序列
作为诱变剂的寡核苷酸序列:
人工合成一段寡核苷酸序列,该序列中 除了所需的碱基突变外,其余的则与目 的基因编码链的特定区段完全互补
Mg2+: 独立作用于每条DNA链,位点随机 Mn2+: 大致在同一位置切割dsDNA
产生平端 或1-2个核苷酸突出
ccc DNA DNase I, Mn2+(low concentration)
Klenow re-ligation
A digestion
作业
利用寡核苷酸进行定点诱变的方法和原 理
基因家族的改组
交错延伸重组 图10-5
以两个以上的有一定同源性的DNA片段 为模板进行PCR反应,通过交换模板机制 实现DNA序列的重新组装。
不需要DNaseI切割DNA,简化了程序。
随机引发重组 图10-6
用一套随机引物与模板配对延伸,产生 不同位点的短DNA片段混合物(含有少 量点突变),以这些短DNA片段为引物 重新组装成全长基因。
大引物pcr进行定点突变的方法

大引物pcr进行定点突变的方法以大引物PCR进行定点突变的方法引言:PCR(聚合酶链式反应)是一种常用的分子生物学技术,可以通过扩增目标DNA序列来获得足够的DNA量。
在基因工程和分子生物学研究中,需要对目标基因进行定点突变,以研究其功能或改变其特性。
大引物PCR是一种常用的方法,可以实现定点突变。
一、大引物PCR的原理大引物PCR是一种基于PCR技术的方法,通过引入带有突变碱基的引物,使扩增产物发生定点突变。
其原理如下:1. 设计引物:根据目标基因的序列,设计一对引物,其中一个引物带有突变碱基,用于引导定点突变。
2. 反应体系:将目标DNA模板、引物、dNTPs(脱氧核苷酸三磷酸盐)、聚合酶和缓冲液混合,构建PCR反应体系。
3. PCR扩增:通过PCR反应,将目标基因的DNA序列扩增出来。
4. 大引物PCR特点:大引物PCR与常规PCR的区别在于,引物的长度较长,通常为30-50个碱基对,以确保引物与目标DNA序列的准确配对并引导定点突变。
二、大引物PCR的步骤大引物PCR通常包括以下步骤:1. 目标基因的DNA提取:从细胞或组织中提取目标基因的DNA。
2. 引物设计:根据目标基因的序列,设计一对引物,其中一个引物带有突变碱基。
3. PCR反应体系的准备:将目标DNA模板、引物、dNTPs、聚合酶和缓冲液按照一定比例混合,构建PCR反应体系。
4. PCR扩增条件的设置:根据引物的特性和目标基因的长度,设置PCR的温度循环条件,包括变性、退火和延伸等步骤。
5. PCR反应:将PCR反应体系放入热循环仪中进行PCR扩增,通常进行30-40个循环。
6. PCR产物的分离:通过凝胶电泳等方法,将PCR扩增产物与模板DNA分离。
7. 定点突变的验证:对PCR扩增产物进行测序,验证是否实现了定点突变。
三、大引物PCR的优势和应用1. 高效性:大引物PCR可以快速、高效地实现定点突变,无需进行繁琐的基因克隆。
基因定点突变全攻略

基因定点突变全攻略基因定点突变全攻略⼀、定点突变的⽬的把⽬的基因上⾯的⼀个碱基换成另外⼀个碱基。
⼆、定点突变的原理定点突变是指通过聚合酶链式反应(PCR)等⽅法向⽬的DNA⽚段(可以是基因组,也可以是质粒)中引⼊所需变化(通常是表征有利⽅向的变化),包括碱基的添加、删除、点突变等。
定点突变能迅速、⾼效的提⾼DNA所表达的⽬的蛋⽩的性状及表征,是基因研究⼯作中⼀种⾮常有⽤的⼿段。
体外定点突变技术是研究蛋⽩质结构和功能之间的复杂关系的有⼒⼯具,也是实验室中改造/优化基因常⽤的⼿段。
蛋⽩质的结构决定其功能,⼆者之间的关系是蛋⽩质组研究的重点之⼀。
对某个已知基因的特定碱基进⾏定点改变、缺失或者插⼊,可以改变对应的氨基酸序列和蛋⽩质结构,对突变基因的表达产物进⾏研究有助于⼈类了解蛋⽩质结构和功能的关系,探讨蛋⽩质的结构/结构域。
⽽利⽤定点突变技术改造基因:⽐如野⽣型的绿⾊荧光蛋⽩(wtGFP)是在紫外光激发下能够发出微弱的绿⾊荧光,经过对其发光结构域的特定氨基酸定点改造,现在的GFP能在可见光的波长范围被激发(吸收区红移),⽽且发光强度⽐原来强上百倍,甚⾄还出现了黄⾊荧光蛋⽩,蓝⾊荧光蛋⽩等等。
定点突变技术的潜在应⽤领域很⼴,⽐如研究蛋⽩质相互作⽤位点的结构、改造酶的不同活性或者动⼒学特性,改造启动⼦或者DNA作⽤元件,提⾼蛋⽩的抗原性或者是稳定性、活性、研究蛋⽩的晶体结构,以及药物研发、基因治疗等等⽅⾯。
通过设计引物,并利⽤PCR将模板扩增出来,然后去掉模板,剩下来的就是我们的PCR 产物,在PCR产物上就已经把这个点变过来了,然后再转化,筛选阳性克隆,再测序确定就⾏了。
三、引物设计原则引物设计的⼀般原则不再重复。
突变引物设计的特殊原则:(1)通常引物长度为25~45 bp,我们建议引物长度为30~35 bp。
⼀般都是以要突变的碱基为中⼼,加上两边的⼀段序列,两边长度⾄少为11-12 bp。
若两边引物太短了,很可能会造成突变实验失败,因为引物⾄少要11-12个bp才能与模板搭上,⽽这种突变PCR要求两边都能与引物搭上,所以两边最好各设⾄少12个bp,并且合成多⼀条反向互补的引物。
基因定点诱变
西北师范大学
基因工程
第二章 基因工程技术原理
返回目录
第四节 基因定点诱变
返回第二章
2.4.1 盒式诱变
利用一段人工合成 的具有突变序列的 寡核苷酸片段取代 野生型基因中的相 应序列.
西北师范大学
2.4.2 寡核苷酸引物诱变
使用化学合成的含有突变碱基 的寡核苷酸片段作引物,启动 单链为了 新合成DNA子链的一个组成 部分。
西北师范大学
2.4.3 PCR诱变 (1) 重组PCR定点诱变: 克服寡核苷酸引物诱 变能力仅限于5’端. 特点:经3轮PCR反应, 一对互补的带有突变 碱基的内侧引物和2个 外侧引物
基因工程
第二章 基因工程技术原理
西北师范大学
(2) 大引物诱变:核心 是第一轮PCR产物为 第二轮PCR引物
基因工程
基因工程
第二章 基因工程技术原理
西北师范大学
寡核苷酸引物诱变过程
正链DNA合成 突变引物合成 异源双链DNA分子制备 闭环异源双链DNA分子富集 转化 突变体筛选 寡核苷酸引物诱变法的局限性
异源双链DNA分子并非真正异源 突变体子代中突变碱基被错配修 复体系修复
基因工程
第二章 基因工程技术原理
西北师范大学
基因工程
第二章 基因工程技术原理
提高寡核苷酸引物诱变突变效率的方法
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
(2) 大引物诱变:核心 是第一轮PCR产物为 第二轮PCR引物
返回目录
返回第二章
返回目录
第四节 基因定点诱变
返回第二章
2.4.1 盒式诱变 利用一段人工合成 的具有突变序列的 寡核苷酸片段取代 野生型基因中的相 应序列.
2.4.2 寡核苷酸引物诱变
使用化学合成的含有突变碱基
的寡核苷酸片段作引物,启动
单链DNA分子进行复制,随
后这段寡核苷酸引物便成为了
新合成DNA子链的一个组成 部分。
寡核苷酸引物诱变过程
正链DNA合成 突变引物合成 异源双链DNA分子制备 闭环异源双链DNA分子富集 转化 突变体筛选 寡核苷酸引物诱变法的局限性
异源双链DNA分子并非真正异源 突变体子代中突变碱基被错配修 复体系修复
提高寡核苷酸引物诱变突变效率的方法 (1) Kunkel 定点诱变法: 1985年由 T.A.Kunkel发明 ung:尿嘧啶脱糖苷酶
(2) 硫代磷酸诱变法: 已知有些限制 酶不能切割硫代磷酸DNA分 子.在异源双链DNA分子制备 时,加入硫代核苷酸,使之掺 入突变链中.然后用前述酶切 割,并用外切酶局部消化后, 进行聚合反应,从而产生具有 定点突变的异源双链DNA
2.4.3 PCR诱变 (1) 重组PCR定点诱 变:克服寡核苷酸引 物诱变能力仅限于5’ 端. 特点:经3轮PCR反 应,一对互补的带有 突变碱基的内侧引 物和2个外侧引物