大豆脱氢抗坏血酸还原酶基因的电子克隆及进化分析
甾醇脱氢酶的基因挖掘、分子改造及其催化合成熊去氧胆酸

甾醇脱氢酶的基因挖掘、分子改造及其催化合成熊去氧胆酸游智能1张仙1李春秀2许建和2(1. 上海百福安生物科技有限公司上海 200030;2. 华东理工大学生物工程学院上海 200237)摘要本文介绍近年来利用以甾醇脱氢酶为核心元件的生物合成法制备熊去氧胆酸的研究进展,并提出了该技术进一步发展所面临的挑战和未来的研究方向,旨在为熊去氧胆酸的生物合成研究提供参考。
关键词熊去氧胆酸甾醇脱氢酶生物合成中图分类号:Q81; Q819 文献标志码:A 文章编号:1006-1533(2024)07-0016-08引用本文 游智能, 张仙, 李春秀, 等. 甾醇脱氢酶的基因挖掘、分子改造及其催化合成熊去氧胆酸[J]. 上海医药, 2024, 45(7): 16-23.Gene mining and molecular modification of hydroxysteroid dehydrogenase and its catalytic synthesis of ursodeoxycholic acidYOU Zhineng1, ZHANG Xian1, LI Chunxiu2, XU Jianhe2(1. Shanghai Bioforany Biotechnology Co., Ltd., Shanghai 200030, China; 2. School of Biotechnology,East China University of Science and Technology, Shanghai 200237, China)ABSTRACT The research progress in the biosynthesis of ursodeoxycholic acid with hydroxysteroid dehydrogenase as the core element in recent years is reviewed, and the challenges and future research directions for further development of this technology are proposeed, aiming to provide reference for the ursodeoxycholic acid biosynthesis.KEY WORDS ursodeoxycholic acid; hydroxysteroid dehydrogenase; biosynthesis熊去氧胆酸(ursodeoxycholic acid, UDCA)是传统名贵中药熊胆粉的主要活性成分,其化学名为3α,7β-二羟基-5β-胆甾烷-24-酸。
植物脱氢抗坏血酸还原酶(DHAR)研究进展

植物脱氢抗坏血酸还原酶(DHAR)研究进展
侯艳霞;张文鑫;杨婧;李小艳;郭艳
【期刊名称】《农业技术与装备》
【年(卷),期】2017(000)002
【摘要】脱氢抗坏血酸还原酶是AsA-GSH循环中能够促进抗坏血酸再生的关键酶.文章从植物脱氢抗坏血酸还原酶的发现、功能及分子生物学等方面的研究进展作一综述,以期为该酶的深入研究及其合理利用提供参考.
【总页数】3页(P28-29,32)
【作者】侯艳霞;张文鑫;杨婧;李小艳;郭艳
【作者单位】山西林业职业技术学院园艺系,山西太原030009;山西林业职业技术学院园艺系,山西太原030009;山西林业职业技术学院园艺系,山西太原030009;山西林业职业技术学院园艺系,山西太原030009;山西林业职业技术学院园艺系,山西太原030009
【正文语种】中文
【中图分类】Q943
【相关文献】
1.苹果脱氢抗坏血酸还原酶(DHAR)基因cDNA片段的克隆及序列分析 [J], 张燕子;马锋旺;张军科;梁东
2.猕猴桃果实L-半乳糖内酯脱氢酶和脱氢抗坏血酸还原酶cDNA片段的克隆与序列分析 [J], 牛歆雨;雷玉山;梁东;马锋旺;王西锐
3.棉花单脱氢抗坏血酸还原酶基因的克隆及原核表达 [J], 钱雯婕;王斐;石峰;葛娟;
李鸿彬
4.玉米脱氢抗坏血酸还原酶基因的克隆和特性研究 [J], 袁进成;马海莲;宋晋辉;宋小青;刘颖慧
5.哈密瓜脱氢抗坏血酸还原酶基因的鉴定及冷胁迫表达分析 [J], 宋文;张琴;宁明;周发科;杨新泉;单春会;唐凤仙
因版权原因,仅展示原文概要,查看原文内容请购买。
【国家自然科学基金】_电子克隆_基金支持热词逐年推荐_【万方软件创新助手】_20140729
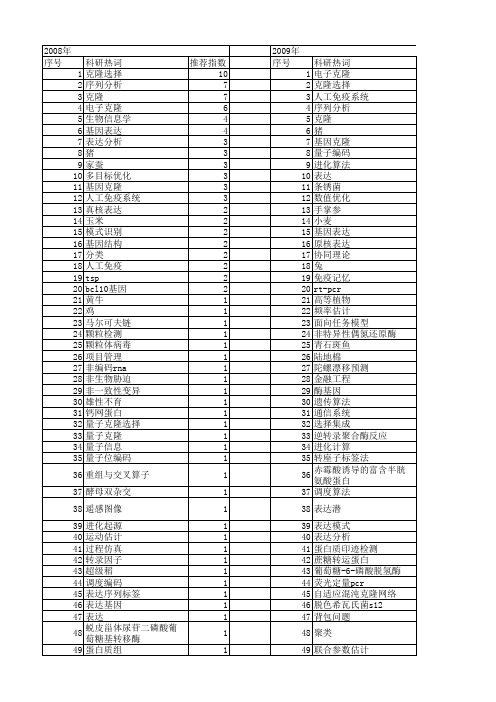
53 54 55 56 57 58 59 60 61 62 63 64 65 66 67 68 69 70 71 72 73 74 75 76 77 78 79 80 81 82 83 84 85 86 87 88 89 90 91 92 93 94 95 96 97 98 99 100 101 102 103 104 105 106
2009年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52
107 108 109 110 111 112 113 114 115 116 117 118 119 120 121 122 123 124 125 126 127 128 129 130 131 132 133 134 135 136 137 138 139 140 141 142 143 144 145 146 147 148 149 150 151 152 153 154 155 156 157 158 159 160
干扰素ε 1 尿苷二磷酸葡萄糖醛酸转移酶 小麦 字符串编码 大鼠 大黄鱼 大豆 多目标 多模式搜索 多执行模式 多年生簇毛麦同 可靠性预测 口蹄疫 受限变异 反转录 原核表达 半随机初始化 半定量rt-pcr 函数优化 入侵性杂草优化 兔 免疫遗传算法 免疫记忆 免疫粒子群 免疫疫苗 免疫克隆算法(ica) 克隆,分子 克降选择 先天免疫 健脾益气 假设检验 信息素 优化 人工免疫算法 人工免疫优化算法 亚组分类 丝氨酸/苏氨酸蛋白磷酸酶 α -醇溶蛋白基因 xenorhabdus xcl1 wrky转录因子 secy sar图像分割 rydber9原子 race pti1
植物脱氢抗坏血酸还原酶(D H A R)研究进展

status of discovery, functions, molecular biology and other aspects of dehydroascorbate reductase was summarized to
provide reference for in-depth study and reasonable utilization of the enzyme.
目前袁发现的 d h ar 基因主要有两种袁分别为 d h ar1 和 d h ar2袁一种存在于胞质中袁另一种为质体酶遥 在同一物种 中袁两种基因的差别主要是其 5爷端核苷酸序列不同袁而不 同物种间同一 d har 基因的 3爷端核苷酸序列不同袁这可能 与基因所在细胞尧物种进化及不同物种的生长环境及对环 境胁迫的响应程度有关遥 邹礼平等[1 1 ]从番茄中获得了这两 种 d h ar 基因袁同时推测 d h ar1 和 d h ar2 分别存在于胞质体 和质体中袁 并证明它们的氨基酸序列仅有 59% 的相似性曰 但他同时发现袁 相同的 d h ar 基因在不同物种间保守性较 高袁番茄 d h ar1 编码的氨基酸序列与水稻尧小麦尧拟南芥尧 烟草中的胞质 d har 基因的氨基酸序列同源性较高袁 均达 到 7 0% 以上曰番茄 d h ar2 编码的氨基酸序列与菠菜尧芥菜尧 青花菜尧 拟南芥中的质体 d h ar 基因的氨基酸序列同源性 也达到了 65% 以上遥 牛歆雨等[1 2]研究也发现袁猕猴桃与苹 果的 dh ar 基因核苷酸序列相似性高达 99% 袁与烟草尧番茄 和土豆的相似性也达到 7 5% 以上遥 可见袁相同来源的 d h ar 基因有较高保守性遥 3 .3 d h a r 基因的表达与调控
28 窑调查研究窑
文章编号院1 6 7 3 - 8 8 7 X (2 0 1 7 )0 2 - 0 0 2 8 - 0 3
抗坏血酸抗肿瘤作用机制的研究进展

始分解,失去一个电荷阴性的电子,形成抗坏血酸自由基
(Asc一),这个活性电子还原一个蛋白质.核心金属离子 (protein.centered metal),称之为过渡态金属(如铜、铁),将 3价铁还原为2价铁离子,在此过程中,会形成具有高度活性 的氧离子。再与细胞外液中的氢离子聚合产生心O:(图2)一
Jun-wen,Email:oujunwen@yahoo.com.cn
as
a
【Abstract】In
cancer
foreign countries,ascorbic acid(vitamin C)has been used
not
complementary drug in
treatments for many years.Although it has
18
图2抗坏lflL酸在细胞外液产生过氧化氧(H。O二)的假设机棚a1
3.2.2.3
H20:更喜欢肿瘤细胞
肿瘤细胞对H20:的易感性考虑与以下几个方面有关: (1)癌细胞自身抗氧化酶(如H20:酶、超氧化物歧化酶)减 少123-241,清除H20:的能力较正常细胞差;(2)细胞内过渡态金 属活性的增强阎;(3)肿瘤细胞以无氧糖酵解代谢为主,需要 大量的葡萄糖提供能量,为此癌细胞会增加GLUT载体的数量 以满足自身能量的需要,而药理浓度的抗坏血酸只能依赖 GLUT转运体进入细胞内,与葡萄糖共同使用GLUT载体转运,
学研究显示,假如健康人体细胞每天受到10'单位的氧化冲
击,就可能导致20余种DNA氧化损伤。体内的修复系统只能 修复99%的DNA损伤,仍有1%的损伤被保留下来。因此,细 胞DNA氧化损伤和突变可随年龄增长而逐步累积。抗坏血酸 在生理学浓度下作为强大的抗氧化剂可以通过清除潜在的诱 导畸变的活性氧簇,防止细胞内DNA的氧化损伤和损伤蓄积。 而且,它还能增强淋巴细胞对氧自由基损伤的防御作用,提高
向日葵抗坏血酸过氧化物酶的电子克隆和生物信息学分析

smi r y w t h to t e p c e n DN a d p o en l v l o s s i g t e d ma n fP r x d s i l i i t a fo h r s e is i A n r ti e e ,p s e sn h o i s o e o i a e a t h
I i c n Cl n n n o n o m a i a y i f n S l o o i g a d Bi i f r tc An l ss o i
As o b t r x d s n n H ein h sa u s c r a e Pe o i a e Ge e i l t u nn u a
大豆GmGST7_基因耐酸铝功能研究

胡康, 金晓雨, 张雪, 等. 大豆GmGST7基因耐酸铝功能研究[J]. 华南农业大学学报, 2023, 44(5): 769-779.HU Kang, JIN Xiaoyu, ZHANG Xue, et al. Study on the tolerant function of soybean GmGST7 gene to acidic aluminum stress[J]. Journal of South China Agricultural University, 2023, 44(5): 769-779.大豆GmGST7基因耐酸铝功能研究胡 康 ,金晓雨,张 雪,王金玉,程艳波,连腾祥,年 海,马启彬(华南农业大学 农学院/国家大豆改良中心广东分中心/亚热带农业生物资源保护与利用国家重点实验室/广东省分子育种重点实验室/岭南现代农业科学与技术广东省实验室, 广东 广州 510642)摘要: 【目的】耐酸铝基因GsMYB7过表达转化大豆品种‘华春6号’后,从转基因株系的表达谱中获得目标基因GmGST7,该基因受酸铝胁迫诱导上调,且位于GsMYB7基因下游,进一步分析其耐酸铝功能,以期提高大豆酸铝耐受能力。
【方法】采用生物信息学方法分析GmGST7基因的碱基序列、蛋白结构域和构建系统进化树。
通过烟草叶片瞬时转化法完成亚细胞定位。
通过RT-qPCR 分析该基因组织表达特异性。
设计0、25、50、75 和 100μmol/L 5个AlCl 3浓度梯度,研究GmGST7对酸铝胁迫的响应。
在50 μmol/L AlCl 3处理下,设计0、4、8、12、16、24、36、48和72 h 共9个时间梯度,对GmGST7的表达模式进行分析。
过表达GmGST7基因遗传转化拟南芥,鉴定阳性植株,并对转基因株系进行耐酸铝表型验证、氧化水平测定、耐酸铝标志基因及下游基因的表达分析。
【结果】GmGST7基因位于大豆第7号染色体,序列全长为1 128 bp 。
条斑紫菜抗坏血酸过氧化酶基因的克隆及生物信息学分析

i n A g r i c u l t u r a l S c i e n c酸过氧化酶基 因的克隆及生物信息学分析
陈吻升 , 席海秀 , 佟少 明, 侯 和 胜
( 辽 宁师 范大 学 生命 科 学学 院 , 辽 宁 大连 1 1 6 0 8 1 )
过程 。
关键词 : 条 斑紫菜 ; 抗坏 血酸过 氧化 物酶 ; 克隆; 生物 信息学
中图分 类号 : ¥ 9 6 8 . 4 3 1 文献 标识码 : A D O I 编码 : 1 0 . 3 9 6 9 / j . i s s n . 1 0 0 6 - 6 5 0 0 . 2 0 1 4 . 0 4 . 0 0 3
植 物在干旱 、 高盐、 低温 、 高 温 等 逆 境 中 产 生 过 量 的活 性 氧 基 团 ( R e a c t i v e o x y g e n s p e c i e s , R O S ) , 细 胞 内 高浓 度 的 R O S会 引 起 细 胞 凋 亡 甚 至 坏 死 。 抗坏血酸过氧化酶 ( A s c o r b a t e p e r o x i d a s e , A P X) 是
h i g h l y h o m o l o g y t o c y t o p l a s m A P X o f h i g h e r p l a n t A r a b i d o p s i s t h a l i a n a , i m p l y i n g P y A P X f u n c t i o n i n g i n c y t o p l a s m . Me a n w h i l e , t h i s
Ab s t r a c t :A n o v e l g e n e o f a s c o r b a t e p e r o x i d a s e wa s c l o n e d f r o m P o r p h y r a y e z o e n s i s , wh i c h wi t h f u l l — l e n g t h o f 8 4 7 b p a n d a n o p e n
维生素C与基因互作的研究

维生素C与基因互作的研究摘要:维生素C(又称抗坏血酸)是一种动物机体所必需的营养素,它是电子供体,因此具有抗氧化性。
本文主要介绍维生素C的性质,维生素C在植物体和某些动物体内的合成途径,以及其在机体内对基因表达的作用,以及近年来对维生素C的研究进展。
关键词:维生素C、抗氧化性、基因表达、合成途径、作用正文:1 维生素C的简介中文名称:维生素C英文名称:vitamin C其他名称:抗坏血酸(ascorbic acid)维生素c键线式:定义:显示抗坏血酸生物活性的化合物的通称,是一种水溶性维生素,水果和蔬菜中含量丰富。
在氧化还原代谢反应中起调节作用,缺乏它可引起坏血病。
维生素C(维生素C),又称抗坏血酸(ascorbic acid, AsA),是普遍存在于植物组织中的高丰度小分子抗氧化物质。
某些动物如人类由于缺乏其合成关键酶L-古洛糖内酯氧化酶不能自身合成维生素C,并且其在人体内不能长久贮存,只能不断从食物中获取维生素C而作为主要维生素C来源的水果和蔬菜其维生素C水平差异较大。
因此维生素C 含量已成为衡量农产品品质的重要指标。
此外,植物中的维生素C 还在抗氧化和清除自由基、光合作用和光保护、细胞的生长和分裂以及参与某些次生代谢物和乙烯的合成等诸多方面起着非常重要的生理功能。
2 植物中维生素c合成的途径2.1 与动物合成途径类似的古洛糖途径Isherwood 等最早提出的类似于动物合成途径的高等植物维生素C生物合成途径,该途径认为植物由D-半乳糖经D-半乳糖醛酸和L-半乳糖内酯(L-GalL)等重要中间物质最终形成维生素C,其间发生了类似动物的碳链倒位。
支持该途径的证据为在植物体内确实存在天然L-GalL并可通过半乳糖内酯脱氢酶(GalLDH)氧化生成维生素C,同时D-半乳糖醛酸(甲酯)也可作为维生素C合成的底物。
但随后的同位素放射性示踪证明植物合成维生素C的过程并未发生碳链的倒位。
虽然有证据显示L-GalL是植物合成维生素C的底物,但D-半乳糖醛酸并不是植物合成维生素C2.2 邻酮醛糖途径为符合同位素放射性示踪实验结果,Loewus等提出的一条非倒位途径即临酮醛糖途径。
高等植物厌氧适应的生理及分子机制

2.4 植物的低氧诱导适应
有许多证据表明,旱生植物的厌氧适应能力受低氧预处理的诱导。植物种间 耐厌氧性的遗传差异乃是植物根系长期适应不同氧分压进化的结果,湿生植 物的这一适应能力强于旱生植物。同一种植物品种间的遗传差异亦有类似的 适应过程。但植物具有潜在的适应厌氧环境的能力,低氧信号能激活某些厌 氧反应基因的表达,使同一种基因型的耐渍能力得到很大的改善。
2.2 代谢适应
2.2.1无氧呼吸的启动
缺氧触发旱生植物的糖酵解过程,首先表现
为乳酸脱氢酶(LDH)活性升高,乳酸发酵。当胞
质中pH值达到6.8时,乙醇脱氢酶(ADH)与丙酮
酸脱羧酶(PDC)被激活,迅速进入乙醇发酵途径。
无氧呼吸是植物潜在的代谢功能,当根细胞中
氧浓度低于某一个阈值--称临界氧分压(COP),
图7 通气组织的形成可以不是被缺氧而是低氧刺激。ACC到乙烯的转化 需要氧的存在。乙烯产生导致根皮层细胞中一系列相关酶厌氧诱导表达, 细胞液泡化,细胞壁微纤丝断裂重接,使中皮层细胞中央贯通,形成大 的通气腔。且周围仍有细胞质流,并不影响离子吸收和运输。
2.1.2 形成不定根
在厌氧下许多植物初生根 受到伤害或死亡,但在靠近地面 处能产生许多新生的不定根。这 一现象乃是由于近地表O2分压较 高所致,也可由乙烯诱导生成, 但不同植物之间的差异很大。不 定根产生的多寡反映了植物的适 应能力,是许多旱生植物中耐渍 基因型的重要标志。
2 植物厌氧适应的策略
2.1 形态适应
2.1.1 通气组织的形成
通气组织(aerenchyma)是植物薄壁组织内一些气腔或空 腔的集合。在厌氧下植物根皮层细胞常发生编程性死亡, 生成通气组织,把氧气运送到根基部,以保证根的代谢需 要,这一过程对于植物在缺氧条件下的生长具有重要的意 义,是植物的一种重要的避缺氧机制。
2014安徽省自然科学基金

项目编号 1408085MC56 1408085MC57 1408085MC58 1408085MC59 1408085MC60 1408085MC61 1408085MC62 1408085MC63 1408085MC64 1408085MC65 1408085MC66 1408085MC67 1408085MC68 1408085MD69 1408085MD70 1408085MD71 1408085MD72 1408085MD73 1408085MD74 1408085MD758085ME78 1408085ME79 1408085ME80 1408085ME81 1408085ME82 1408085ME83 1408085ME84 1408085ME85
主持人 周艺峰 凤权 朱曙光 朱大焕 梁凤霞 余庆选 陶锋 王伟 刘有余 胡毅 丁卫平 乔红斌 黎春林 宋新江 张晶晶 胡业林 朱茂飞 吕小莲 李家文 郭兴众 刘广彬 陈清华 贾磊 来永斌 葛斌 朱家兵 王子磊 方贤勇 董永平 史明光
依托单位 安徽大学 安徽工程大学 安徽建筑大学 中国科学院合肥物质 科学研究院 合肥工业大学 中国科学技术大学 安徽工程大学 合肥工业大学 安徽工程大学 合肥工业大学 中国科学技术大学 安徽工业大学 铜陵学院 安徽省水利部淮河水 利委员会水利科学研 合肥工业大学 安徽理工大学 中国科学院合肥物质 科学研究院 滁州学院 中国科学技术大学 安徽工程大学 合肥通用机械研究院 安徽理工大学 合肥通用机械研究院 安徽理工大学 安徽理工大学 中国电子科技集团第 三十八研究所 中国科学技术大学 安徽大学 安徽工业大学 合肥工业大学
附件
2014年度安徽省自然科学基金项目表
一、2014年度面上项目
项目编号 1408085MA01 1408085MA02 1408085MA03 1408085MA04 1408085MA05 1408085MA06 1408085MA07 1408085MA08 1408085MA09 1408085MA10 1408085MA11 1408085MA12 1408085MA13 1408085MA14 1408085MA15 1408085MA16 1408085MA17 1408085MA18 1408085MA19 1408085MA20 1408085MA21 1408085MB22 1408085MB23 1408085MB24 1408085MB25 主持人 黄守军 王奇 周兴才 汪忠志 钱建发 程燕 方龙祥 叶永升 申传胜 张榕京 鲁文建 张战军 朱青峰 郑云英 王大理 李大创 郭玉献 喻远琴 黄仙山 张刚 余龙宝 裘灵光 江海龙 谷永红 汪文栋 依托单位 安徽师范大学 安徽大学 铜陵学院 安徽工业大学 安徽理工大学 中国人民解放军陆军 军官学院 安徽师范大学 淮北师范大学 安庆师范学院 中国科学技术大学 中国科学院合肥物质 科学研究院 安徽大学 中国科学技术大学 淮北师范大学 安徽师范大学 合肥师范学院 安徽建筑大学 安徽大学 安徽工业大学 皖西学院 合肥师范学院 安徽大学 中国科学技术大学 中国科学技术大学 中国科学技术大学 项目名称 弹性材料若干数学问题的研究 脉冲微分方程若干问题的理论与应用研究 复杂相依数据半参数模型非线性小波估计的理论研究及其应用 信源广义熵遍历定理及其应用基础研究 量子编码理论中新问题的研究与探索 一类非线性时滞微分方程奇异摄动问题研究 无限二阶矩过程驱动的OU过程的统计推断 图的pebbling问题和Graham猜想 复杂网络上多尺度动力学过程的介观理论研究 微生物膜(Biofilm)的成因,发展及演化 过渡金属硫族化合物中电荷密度波与超导电性的关联性研究 量子关联研究方法发展与量子态关联研究 大质量恒星形成区谱线观测研究 粘弹性多孔介质的分数阶建模及有限元分析 受限Dirac电子系统中奇异的量子效应研究 一类特殊的固态海森堡系统中的复杂量子特性研究 二氧化钒微纳尺寸晶粒的相变特性及调控机理 醇-水溶液团簇结构中蓝移氢键C-H„O的理论与实验研究 白光腔实现可调短脉冲激光器的机理研究 基于几何量子操作的固态量子计算研究 基于超导线路的自旋-玻色模型理论实现及其性质研究 基于金属-有机骨架纳米晶荧光试纸的制备及其对硝基爆炸物传感性 质研究 催化功能导向金属有机框架材料的结构调控与性能研究 手性烯丙醇的取代反应 可控形貌钛酸前体制备TiO2复合纳米结构及其催化性能研究
野生大豆P5CS基因的克隆及对盐胁迫反应

植物遗传资源学报2014,15(4):844⁃849JournalofPlantGeneticResourcesDOI:10.13430/j.cnki.jpgr.2014.04.024野生大豆P5CS基因的克隆及对盐胁迫反应张兆元1,陈吉宝1,宋佳璘2,曹苑楠1,冉小芹1,党虹1(1南阳师范学院,河南南阳473061;2河南省环境监控中心,郑州450004)㊀㊀摘要:逆境下植物大量积累脯氨酸是减轻胁迫伤害的一种自我保护机制㊂本研究应用同源克隆方法从NaCl处理的野生大豆中克隆获得一个脯氨酸合成酶(P5CS)基因,命名为GsP5CS㊂该基因核苷酸序列全长2 232kb,含一个2148bp开放阅读框,编码715个氨基酸,包含有高等植物P5CS蛋白质的5个主要功能域,与菜豆PvP5CS1基因核苷酸序列相似性高达98 79%㊂RealTimePCR分析显示该基因受轻度盐胁迫诱导上调表达,根中表达高峰出现在200mmol/LNaCl处理下,相对表达量为对照的5 83倍;叶片中表达高峰出现在300mmol/LNaCl处理条件下,相对表达量为对照的12 78倍㊂并且该基因在根和叶片中的表达模式和脯氨酸含量的变化模式相同㊂上述结果说明,GsP5CS可能参与野生大豆脯氨酸合成㊂㊀㊀关键词:野生大豆;P5CS基因;盐胁迫;表达模式;RealTimePCRCloningandExpressionAnalysisofP5CSGenefromWildSoybean(Glycinesoja)SeedlingunderSaltStressZHANGZhao⁃Yuan1,CHENJi⁃Bao1,SONGJia⁃Lin2,CAOYuan⁃Nan1,RANXiao⁃Qin1,DANGHong1(1NanyangNormalUniversity,NanyangHenan473061;2EnvironmentalMonitoringCenterinHenanProvince,Zhengzhou450004)Abstract:Prolineaccumulationinplanttissuesisaself⁃protectionmechanismreducingstressinjuries.Inthestudy,aP5CSgenewasisolatedbyhomologousclonestrategyfromwildsoybeanseedingunderNaCltreatment,namedGsP5CS.Sequenceanalysisshowedthatthefull⁃lengthcDNAwas2 232kb,containinganopenreadingframeof2148bp,andencoding715aminoacidswith5typicalfunctiondomainsofplantP5CSprotein.ThecDNAsequenceshared98 79%similaritywithPvP5CS1fromcommonbean.RealtimePCRanalysisshowedthatthegeneexpressionlevelwassignificantlyup⁃regulatedundersaltstress.Therelativeexpressionofthegenereachedtothemaximumunder200mmol/LNaCltreatmentinrootsand300mmol/LNaCltreatmentinleavesandwerere⁃spectively5 83and12 78timesofthecontrol.AndtheexpressionmodelsofGsP5CSgeneweresimilarwiththeprolineaccumulationmodelsinleaveandrootofwildsoybeantreatedwithdifferentNaClconcentration.TheaboveresultsshowedthatGsP5CSenzymemightparticipateintheprolinesynthetaseinwildsoybean(Glycinesoja).Keywords:wildsoybean;P5CSgene;saltstress;expresspattern;realtimePCR收稿日期:2013⁃10⁃21㊀㊀㊀㊀修回日期:2013⁃11⁃14㊀㊀网络出版日期:2014⁃06⁃09URL:http://www.cnki.net/kcms/detail/11.4996.S.20140609.1405.002.html基金项目:南阳师范学院专项项目(ZX2010008);河南省教育厅自然科学研究项目(2010A180016)第一作者主要从事植物基因工程研究㊂E⁃mail:ying0482@qq.com通信作者:陈吉宝,主要从事植物生物化学与分子生物学研究㊂E⁃mail:chenjibao2012@163.com野生大豆是栽培大豆的近缘野生种,是大豆育种极为重要的种质资源㊂野生大豆的研究利用,对拓宽栽培大豆的遗传基础,提高大豆育种水平有重要价值[1]㊂前人的研究证明野生大豆和大豆都具有很强的抗盐能力[2⁃4],但是野生大豆耐盐性相关的理论研究至今尚未有突破性进展㊂非生物逆境胁迫是作物产量降低的主要原因之一,脯氨酸是植物细胞中的渗透调节物质,在渗透胁迫条件下脯氨酸的积累不仅有助于提高植物细胞的渗透势,提高植物抗胁迫能力,而且还是植物从胁迫条件恢复正常过程中迅速有效的氮源㊁碳源和还原剂[5⁃6],因此提高植物逆境下脯氨酸的积累速度对于提高植物抗渗透胁迫能力具有重要的意义㊂脯氨酸合成酶Δ⁃吡咯啉⁃5⁃羧酸合成酶(P5CS,pyrroline⁃5⁃carboxylate㊀4期张兆元等:野生大豆P5CS基因的克隆及对盐胁迫反应synthetase)是植物脯氨酸生物合成中的限速酶,决定着植物体内脯氨酸的积累速度[7]㊂目前很多植物P5CS基因已获得较深入的研究[8⁃12],但对野生大豆脯氨酸合成酶基因仍然没有很好的研究,本研究通过克隆野生大豆脯氨酸合成酶基因并分析其对盐胁迫的反应,旨在从分子生物学角度解释野生大豆抗盐机理㊂1㊀材料与方法1 1㊀野生大豆幼苗培养野生大豆种子用98%硫酸处理10min,无菌水清洗数遍至pH值接近中性后用无菌水室温培养至发芽㊂将发芽的种子单粒点播于营养钵内,营养土用花土ʒ细砂ʒ壤土按照1ʒ1ʒ1混合配制,每钵使用50g营养土㊂营养钵置于室温光照16h/d培养至第1对真叶完全展开,培养期间每隔3d用自来水浇灌营养土㊂在第1对真叶完全展开第2对真叶刚刚露出时对所有幼苗进行一次充分灌溉,然后自然干旱控水,5d后选择大小一致㊁健康无损的幼苗进行盐胁迫处理㊂每钵分别用100mL浓度为100mmol/L㊁200mmol/L㊁300mmol/L㊁400mmol/L㊁500mmol/LNaCl溶液均匀浇灌营养土5h模拟盐胁迫处理㊂每个处理重复6次,以自来水浇灌营养土为对照㊂1 2㊀总RNA提取及cDNA第1链合成分别取盐处理野生大豆幼苗叶片㊁茎和根,液氮速冻后用Trizol试剂提取总RNA㊂按照TaKaRa公司的PrimeScript2试剂盒说明,用Oligo(dT)15作为引物合成cDNA第1链㊂反转录产物稀释10倍备用㊂1 3㊀目的基因的扩增参考普通菜豆P5CS基因序列(GeneBank:EU407263),利用DNAStar软件包中的PrimerSelect软件设计引物对G1:5ᶄ⁃ATGGAGAACACAGATCCT⁃TG⁃3ᶄ,G2:5ᶄCTAGAGGAAATAAAACACAGGCT⁃3ᶄ,由上海生工生物工程技术有限公司合成㊂PCR反应体系总体积20μL,包括反转录产物4μL作模板,目标基因上下游引物(引物浓度2μmol/L)各3μL㊁2ˑTaqPCRMasterMix10μL㊂PCR扩增程序为95ħ预变性3min;95ħ变性30s,50ħ退火40s,72ħ延伸1min30s,33个循环,72ħ延伸10min㊂1 4㊀目标基因表达量实时荧光定量PCR检测按照RealMasterMix(SYBRGreen)PCR试剂盒(TIANGENCorporation)操作,以大豆β⁃Actin基因作为内参,采用实时荧光定量PCR(RT⁃qPCR,real⁃timequantitativePCR)检测基因相对表达量㊂目标基因引物对为F:5ᶄ⁃GGCCCTCCGAGTGACCAA⁃3ᶄ和R:5ᶄ⁃CTCGCGGGCGTTAACCTCTTTT⁃3ᶄ㊂RT⁃qPCR反应程序为:94ħ预变性2min;94ħ变性20s,60ħ退火30s,72ħ延伸40s,40个循环,72ħ延伸10min㊂基因的表达量采用K.J.Livak等[13]的2-ΔΔCT公式计算,其中ΔΔCT=CT,Target-CT,Actin㊂CT,Target和CT,Actin分别是目标基因和内参基因在不同处理点的CT值,每个样品3次重复㊂1 5㊀序列分析目标扩增产物由上海英骏生物技术有限公司进行测序,采用DNAStar软件系统的Editsep㊁MegAlign软件包以及DNAMAN软件分析基因序列㊂1 6㊀脯氨酸含量检测取盐胁迫处理幼苗真叶和根在液氮中充分研磨后,参照陈红敏等[14]的方法分别测定脯氨酸含量㊂2㊀结果与分析2 1㊀RT⁃PCR扩增目的基因用目标基因特异引物对G1/G2,以野生大豆幼苗叶片反转录产物为模板进行PCR扩增,扩增产物电泳检测结果显示获得一条约2 2kb左右的扩增片段,大小与目标片段一致(图1)㊂测序结果表明,该片段全长2 232kb,有完整的开放阅读框,无5ᶄUTR序列,3ᶄUTR序列长84bp,编码715个氨基酸,蛋白质的大小约77 42kD(图2),将其命名为GsP5CS㊂图1㊀GsP5CS基因RT⁃PCR扩增结果Fig 1㊀RT⁃PCRproductofGsP5CSgene2 2㊀野生大豆P5CS基因序列分析张春宝等[19]2007年曾报道2个P5CS基因,分别为GsP5CS1和GsP5CS2,其序列全长分别为1568bp和560bp㊂序列比对表明,GsP5CS1与GsP5CS2的核苷酸相似性为72 7%,本研究获得GsP5CS和GsP5CS1㊁GsP5CS2的核苷酸相似性分别548植㊀物㊀遗㊀传㊀资㊀源㊀学㊀报14卷图2㊀野生大豆P5CS基因序列及蛋白质序列Fig 2㊀NucleotidesequencesandputativeaminoacidssequencesofGsP5CSgene为89 3%和74 1%,说明GsP5CS可能和GsP5CS1是同一个基因㊂GsP5CS1第1个核苷酸起始于GsP5CS的第580bp处,其末端序列结束于GsP5CS的第2148bp㊂从GsP5CS蛋白的氨基酸序列比对结果可以看出,GsP5CS的起始氨基酸和大豆㊁豇豆㊁普通菜豆㊁拟南芥以及小麦P5CS蛋白的起始氨基酸完全相同(图3),因此认为GsP5CS具有完整的开放阅读框,而GsP5CS1不具有完整的开放阅读框㊂利用DNAStar软件系统中的MegAlign软件包,分析野生大豆P5CS基因与其他植物P5CS基因序列的多态性,结果表明GsP5CS与大豆GmP5CS(AY492005)㊁菜豆PvP5CS1(EU340347)和PvP5CS2(EU407263)㊁豇豆VaP5CS(M92276)㊁拟南芥AtP5CS1(X86777)和AtP5CS2(Y09355)㊁小麦TaP5CS(AY888045)基因的核酸序列相似性分别为93 6%㊁83 3%㊁98 79%㊁83 2%㊁73 1%㊁73 3%㊁69 4%,氨基酸序列的相似性分别为93 2%㊁84 4%㊁97 9%㊁82 1%㊁75 6%㊁74 7%㊁73 9%㊂采用ClustalW程序分析了GsP5CS与其他P5CS基因的进化关系,结果显示野生大豆与菜豆PvP5CS2亲缘关系最近,其次是大豆GmP5CS,亲源关系最远的是单子叶植物小麦(图4)㊂648㊀4期张兆元等:野生大豆P5CS基因的克隆及对盐胁迫反应下划线部分表示5个主要功能域的序列Sequencesunderlinedrepresenttheconservedfunctiondomains图3㊀GsP5CS基因与几种植物P5CS基因的氨基酸序列比对Fig 3㊀AminoacidsequencealignmentofGsP5CS1withP5CShomologsinplants图4㊀P5CS基因的系统进化树Fig 4㊀PhylogenetictreeofP5CSgene2 3㊀盐胁迫下野生大豆幼苗脯氨酸含量的变化在整个处理期间,叶片和根中脯氨酸水平呈现先升后降的变化趋势(图5)㊂叶片中脯氨酸从21 57μg/g㊃FW(对照)逐渐上升到最大值64 59μg/g㊃FW(300mmol/L),为对照的2 9倍,然后又逐渐下降到44 96μg/g㊃FW(500mmol/L)㊂方差分析显示,叶片中脯氨酸含量在不同胁迫点之间差异极显著(P<0 01)㊂根中脯氨酸从18 12μg/g㊃FW(对照)逐渐上升到最大值39 12μg/g㊃FW(200mmol/L),为对照的2 1倍,然后又逐渐下降到24 45μg/g㊃FW(500mmol/L)㊂方差分析显示,根中脯氨酸含量在200mmol/L和300mmol/L之间差异不显著,对照和其他各胁迫点之间差异极显著(P<0 01)㊂盐胁迫下叶中脯氨酸积累量都极显著高于根部(P<0 01),最高可达根部的2 0倍(400mmol/L)㊂2 4㊀野生大豆P5CS基因对盐胁迫的反应应用实时定量PCR的方法分析了盐胁迫处理下GsP5CS基因的相对表达量,结果显示,在整个处理期间,叶片和根中GsP5CS基因的相对表达量也呈现先升后降的变化趋势(图6)㊂无盐胁迫(对照)748植㊀物㊀遗㊀传㊀资㊀源㊀学㊀报14卷图5㊀NaCl胁迫下野生大豆幼苗叶片和根中脯氨酸含量Fig 5㊀ProlinecontentinleavesandrootsofwildsoybeanseedlingstreatedwithNaClstress情况下,GsP5CS基因在根中和叶片中的相对表达量无显著差异㊂轻微的盐胁迫(100mmol/LNaCl)即可诱导叶片和根中GsP5CS基因的上调表达,其相对表达量分别是对照的1 78倍和4 63倍㊂在200mmol/LNaCl处理下根中GsP5CS基因相对表达量就达到最大值,为对照的5 83倍;叶片中GsP5CS基因最大相对表达量出现在300mmol/LNaCl处理条件下,为对照的12 78倍㊂在500mmol/LNaCl处理下叶片中GsP5CS基因相对表达量极显著小于对照(P<0 01),为对照的0 28倍,虽然根中基因的相对表达量也有所下降,但比对照还略高,为对照的1 28倍㊂图6㊀NaCl胁迫下野生大豆幼苗叶片和根中GsP5CS基因mRNA的相对表达量Fig 6㊀GsP5CSgenemRNArelativelevelsinleavesandrootsofwildsoybeanseedlingstreatedwithNaClstress3㊀讨论植物体内脯氨酸的合成有2条途径,即谷氨酸途径和鸟氨酸途径,其中谷氨酸途径被认为是植物逆境胁迫相关途径㊂在谷氨酸途径中,脯氨酸的合成起始于谷氨酸,在Δ⁃吡咯啉⁃5⁃羧酸合成酶(P5CS)的催化下转化成谷氨酸半醛,该物质自动转化成吡咯啉⁃5⁃羧酸(P5C,pyrroline⁃5⁃carboxylate),P5C再由吡咯啉⁃5⁃羧酸还原酶(P5CR,pyrroline⁃5⁃carboxylatereductase)催化生成脯氨酸[8,15]㊂P5CS是植物细胞内脯氨酸合成途径的关键酶,目前已从许多植物中分离到P5CS基因,并从分子遗传角度证明该基因是一个有益的植物抗逆境胁迫基因[16⁃17]㊂本研究表明,GsP5CS具有1个2148bp的完整开放阅读框,推定其编码715个氨基酸,与其他物种已证明的P5CS蛋白质相似㊂序列分析结果表明,GsP5CS与同为豆科的PvP5CS2及大豆的GmP5CS同源性最高,与拟南芥㊁小麦的P5CS基因同源性也较高,这与植物分类学上的亲缘关系一致㊂A.C.Turchetto⁃Zolet等[18]通过对48种植物和微生物P5CS基因序列比对分析表明,植物P5CS基因在进化过程发生过基因拷贝数复制现象,即P5CS在植物体内都存在双拷贝㊂张春宝等[19]曾报道了一个野生大豆P5CS基因,本研究又从野生大豆中克隆获得一个P5CS基因,说明野生大豆P5CS基因在进化过程中也出现过基因复制现象㊂本研究结果表明,盐胁迫能够强烈诱导GsP5CS基因的表达,并且脯氨酸含量的变化与GsP5CS的表达量平行,进一步证明GsP5CS基因与野生大豆体内脯氨酸的合成具有高度相关性㊂前人的研究结果证明P5CS基因在干旱㊁高盐等逆境胁迫下大量表达,并调控内源脯氨酸的合成[7,20⁃21],本研究结果与前人的结果类似,说明GsP5CS基因是一个盐胁迫诱导上调表达基因㊂本研究首次发现野生大豆P5CS基因在根和叶片中具有不同的表达规律,在相同胁迫条件下根和叶中该基因的表达有明显的不同,在100mmol/LNaCl处理下,根中GsP5CS基因的表达是叶中的2 6倍,然后随着胁迫程度的加强该基因在叶中的表达量明显高于根部,最高可达根部表达量的4倍(300mmol/LNaCl),同时该基因在根部表达峰值的出现早于叶部㊂以上结果说明该基因在根部对盐胁迫的反应速度快于叶部,但反应强度小于叶部㊂胁迫条件下基因在叶部总相对表达量明显高于根部5CS,这刚好与脯氨酸在根和叶中的积累量不同相吻合,这些结果暗示叶部GsP5CS基因的诱导上调表达速度对提高野生大豆抗盐能力具有积极的意义㊂本研究克隆了一个新的GsP5CS基因,为进一步研究GsP5CS基因表达的调控和野生大豆抗逆分子机理及创造耐盐新品种奠定了基础㊂848㊀4期张兆元等:野生大豆P5CS基因的克隆及对盐胁迫反应致谢:本研究所用野生大豆种子由中国农业科学院作物科学研究所野生大豆课题组王克晶老师提供,特此致谢!参考文献[1]㊀王克晶,李向华.国家基因库野生大豆(Glycinesoja)资源最近十年考察与研究[J].植物遗传资源学报,2012,13(4):507⁃514[2]㊀孙微,张辉.河南和山西野生大豆耐盐鉴定及耐盐相关基因分析[D].北京:中国农业科学院,2007[3]㊀肖鑫辉,李向华,刘洋,等.野生大豆(Glycinesoja)耐高盐碱土壤种质的鉴定与评价[J].植物遗传资源学报,2009,10(3):392⁃398[4]㊀姜静涵,郭勇,常汝镇,等.大豆苗期耐盐性的简便鉴定方法[J].作物学报,2013,39(7):1248⁃1256[5]㊀AkpJnarBA,LucasSJ,BudakH.Genomicsapproachesforcropimprovementagainstabioticstress[J].ScientificWorldJournal,2013,doi./10 1155/2013/361921[6]㊀IsabelPA,FranciscoCA,AnaIM,etal.Pyrroline⁃5⁃carboxylatesynthaseandprolinebiosynthesis:fromosmotolerancetoraremet⁃abolicdisease[J].ProteinSci,2010,19:372⁃382[7]㊀LehmannS,FunckD,SzabadosL,etal.Prolinemetabolismandtransportinplantdevelopment[J].AminoAcids,2010,39:949⁃962[8]㊀陈吉宝,景蕊莲,毛新国,等.转PvP5CS2基因烟草对干旱胁迫的反应[J].植物遗传资源学报,2008,9(2):186⁃189[9]㊀周精华,邢虎成,揭雨成,等.苎麻Δ1⁃吡咯啉⁃5⁃羧酸合成酶(P5CS)基因的克隆和表达分析[J].作物学报,2012,38(3):549⁃555[10]㊀ChenJB,ZhangXY,JingRL,etal.Cloningandgeneticdiversi⁃tyanalysisofanewP5CSgenefromcommonbean(PhaseolusvulgarisL.)[J].TheorApplGenet,2010,120:1393⁃1404[11]㊀KarthikeyanA,PandianSK,RameshM.Transgenicindicaricecv ADT43expressingapyrroline⁃5⁃carboxylatesynthetase(P5CS)genefromVignaaconitifoliademonstratessalttolerance[J].PlantCellTissOrganCult,2011,107:383⁃395[12]㊀曹丽,义鸣放,孙振元,等.多年生黑麦草P5CS基因的定点突变及其在拟南芥中的转化[J].草业学报,2011(1):143⁃152[13]㊀LivakKJ,SchmittgenTD.AnalysisofrelativegeneexpressiondatausingReal⁃TimequantitativePCRandthe2⁃ΔΔCTmethod[J].Methods,2001,25:402⁃408[14]㊀陈红敏,陈明,魏安智,等.抗逆相关基因GmAREB转基因小麦的获得与鉴定[J].植物遗传资源学报,2010,11(6):749⁃754[15]㊀ChakrabortyK,SairamRK,BhattacharyaRC.Salinity⁃inducedexpressionofpyrrolline⁃5⁃carboxylatesynthetasedeterminesalinitytoleranceinBrassicaspp[J].ActaPhysiolPlant,2012,doi:10 1007/s11738⁃012⁃0994⁃y[16]㊀KiranKumarGS,SujataKG,VijayKumarBMR,etal.Heterol⁃ogousexpressionofP5CSgeneinchickpeaenhancessalttoler⁃ancewithoutaffectingyield[J].BiolPlantarum,2011,55(4):634⁃640[17]㊀BagdiDL,ShawBP.AnalysisofprolinemetabolicenzymesinOryzasativaunderNaClstress[J].JEnvironBiol,2013,34:677⁃681[18]㊀Turchetto⁃ZoletAC,Margis⁃PinheiroM,MargisM.Theevolutionofpyrroline⁃5⁃carboxylatesynthaseinplants:akeyenzymeinpro⁃linesynthesis[J].MolGenetGenomics,2009,281:87⁃97[19]㊀张春宝,赵洪锟,李启云,等.野生大豆Δ⁃吡咯啉⁃5⁃羧酸合成酶(P5CS)基因的克隆与序列分析[J].大豆科学,2008,6(27):915⁃920[20]㊀JaarsmaR,RozemarijnSMV,AlbertusHB.Effectofsaltstressongrowth,Na+accumulationandprolinemetabolisminpotato(Solanumtuberosum)cultivars[J].PLoSONE,2013,8(3):1⁃10[21]㊀HuangZ,ZhaoL,ChenD,etal.Saltstressencouragesprolineac⁃cumulationbyregulatingprolinebiosynthesisanddegradationinJerusalemArtichokeplantlets[J].PLoSONE,2013,8(4):1⁃10948。
【国家自然科学基金】_脱氢抗坏血酸还原酶_基金支持热词逐年推荐_【万方软件创新助手】_20140731
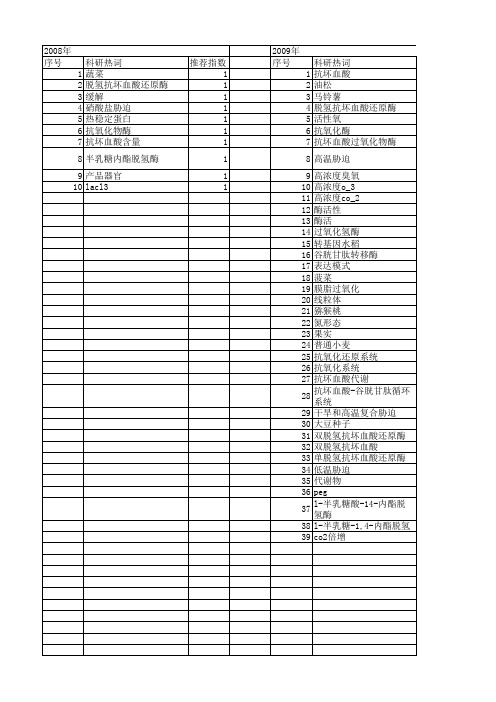
科研热词 蔬菜 脱氢抗坏血酸还原酶 缓解 硝酸盐胁迫 热稳定蛋白 抗氧化物酶 抗坏血酸含量 半乳糖内酯脱氢酶 产品器官 lacl3
推荐指数 1 1 1 1 1 1 1 1 1 1
2009年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39
209 10 11 12 13 14 15 16 17
2011年 科研热词 脱氢抗坏血酸还原酶 马铃薯 酶活 遗传转化 转基因 蛋白表达 臭氧胁迫 温度胁迫 活性氧 江南卷柏 植物表达载体构建 棉花 抗坏血酸 大豆 克隆 gdp-甘露糖焦磷酸化酶 asa-gsh循环 推荐指数 2 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
2013年 序号 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17
科研热词 推荐指数 抗坏血酸-谷胱甘肽循环 2 黄瓜 1 镉 1 铜胁迫 1 谷胱甘肽 1 调节 1 臭氧 1 胁迫 1 硫 1 番茄 1 植物络合素 1 植物 1 抗氧化系统 1 抗坏血酸 1 小白菜 1 土壤紧实胁迫 1 一氧化氮 1
科研热词 抗坏血酸 油松 马铃薯 脱氢抗坏血酸还原酶 活性氧 抗氧化酶 抗坏血酸过氧化物酶 高温胁迫 高浓度臭氧 高浓度o_3 高浓度co_2 酶活性 酶活 过氧化氢酶 转基因水稻 谷胱甘肽转移酶 表达模式 菠菜 膜脂过氧化 线粒体 猕猴桃 氮形态 果实 普通小麦 抗氧化还原系统 抗氧化系统 抗坏血酸代谢 抗坏血酸-谷胱甘肽循环系统 干旱和高温复合胁迫 大豆种子 双脱氢抗坏血酸还原酶 双脱氢抗坏血酸 单脱氢抗坏血酸还原酶 低温胁迫 代谢物 peg l-半乳糖酸-14-内酯脱氢酶 l-半乳糖-1,4-内酯脱氢酶 co2倍增
植物抗坏血酸过氧化物酶的作用机制、酶学及分子特性

植物抗坏血酸过氧化物酶的作用机制、酶学及分子特性一、本文概述植物抗坏血酸过氧化物酶(Ascorbate Peroxidase,AP)是一种在植物细胞内广泛存在的关键酶,其在植物抗氧化防御系统中发挥着至关重要的作用。
本文旨在全面探讨植物抗坏血酸过氧化物酶的作用机制、酶学特性以及分子特性,以期为深入理解植物抗氧化防御系统的运行规律,以及提高植物抗逆性和农业生产力提供理论基础。
我们将详细介绍抗坏血酸过氧化物酶的生化功能,包括其催化抗坏血酸清除活性氧的能力及其在细胞氧化还原稳态中的作用。
接着,我们将深入探讨抗坏血酸过氧化物酶的酶学性质,如酶的动力学特性、抑制剂和激活剂的影响等。
我们将对抗坏血酸过氧化物酶的分子特性进行阐述,包括其基因结构、表达调控以及蛋白质结构等方面的研究。
通过本文的综述,我们期望能够为植物生物学、农业生物技术以及植物抗逆性研究等领域提供有益的参考和启示。
二、植物抗坏血酸过氧化物酶的作用机制植物抗坏血酸过氧化物酶(AP)是一种关键的抗氧化酶,主要作用是清除植物细胞中的过氧化氢(H2O2),以防止氧化应激对细胞造成的损伤。
AP的作用机制主要涉及到酶的催化活性以及其与底物的相互作用。
在AP的催化过程中,抗坏血酸(AsA)作为还原剂,将H2O2还原为水(H2O),而自身则被氧化为单脱氢抗坏血酸(DHA)。
这个过程可以表示为:2AsA + H2O2 → 2DHA + 2H2O。
DHA随后通过抗坏血酸再生系统被还原回AsA,从而维持了AP的催化循环。
AP的作用机制还涉及到其在细胞内的定位。
在植物细胞中,AP 主要分布在叶绿体、细胞质和线粒体等细胞器中。
这些细胞器中的AP通过特定的信号肽序列被定位到相应的位置,从而实现了对特定区域H2O2的高效清除。
AP的活性还受到多种因素的调节,包括光照、温度、pH值以及底物和抑制剂的浓度等。
光照和温度可以影响AP的稳定性和活性,而pH值则可以影响AP与底物的结合能力。
单脱氢抗坏血酸还原酶

单脱氢抗坏血酸还原酶(MDHAR)检测
单脱氢抗坏血酸还原酶(Monodehydroascorbate reductase, MDHAR)是抗坏血酸-谷胱甘肽循环(AsA-GSH)中的一种重要酶,催化单脱氢抗坏血酸(MDHA)还原生成抗坏血酸,在抗坏血酸氧化还原代谢中具有重要作用。
测定原理:MDHAR催化NADH还原MDHA生成抗坏血酸和NAD+,NADH在340nm有特征吸收峰,但是NAD+没有。
通过测定340nm 光吸收下降速率,来计算出单脱氢抗坏血酸还原酶活性。
迪信泰检测平台采用生化法,结合相应的酶类的试剂盒,可高效、精准的检测单脱氢抗坏血酸还原酶的活性变化。
此外,我们还提供其他ASA-GSH循环类的检测服务,以满足您的不同需求。
生化法测定单脱氢抗坏血酸还原酶样本要求:
1. 请确保样本量大于0.2g或者0.2mL。
周期:2~3周
项目结束后迪信泰检测平台将会提供详细中英文双语技术报告,报告包括:
1. 实验步骤(中英文)
2. 相关参数(中英文)
3. 图片
4. 原始数据
5. 单脱氢抗坏血酸还原酶活性信息。