基因克隆的酶学基础2014
第二章基因工程的酶学基础

经同尾酶消化的DNA末端连接示意图
BamH I 5' XXXXGGATCCXXXXXX 3' 3' XXXXCCTAGGXXXXXX 5'
Bgl II 5' XXXXAGATCTXXXXXX 3' 3' XXXXTCTAGAXXXXXX 5'
5' XXXXG GATCCXXXXXX 3' 3' XXXXCCTAG GXXXXXX 5'
限制性核酸内切酶的分类
I型
II 型
III 型
1. 限制修饰活性 单一多功能的酶 限制酶和修饰酶分开 双功能酶
2. 内切酶的蛋白 3种不同亚基
单一成分
2种亚基
质结构
3. 限制辅助因子 ATP、Mg2+和S- Mg2+ 腺苷甲硫氨酸
ATP、Mg2+和S腺苷甲硫氨酸
4. 切割位点
5. 特异性切割 6. 基因克隆中
距特异性位点 1000bp 不是
无用
特异性位点及其附近 特异性位点3‘端 24-26bp处
是
是
非常有用
有用
限制性核酸内切酶的命名
属名
种名
株名
Haemophilus influenzae d 嗜血流感杆菌 d株
H i n d III H i n d III
同一菌株中所含的多个不同的限制性核酸内切酶
第一节 限制性核酸内切酶
4×10 — 4 1
大肠杆菌B
10 — 4
大肠杆菌K
1 E.O.P 成斑率
efficiency of plating
50年代初发现了由寄主控制的限制和修饰现象
基因克隆简介ppt课件

5’3’-
5’3’-
GAATTC CTTAAG
G AATTC CTTAA G
-3’ -5’
-3’ -5’
14
(4)粘性末端的意义 ①连接便利
i)不同的DNA双链: 只要粘性末端碱基互补就可以连接。 这比连接两个平齐末端容易的多。
ii)同一个DNA分子内连接: 通过两个相同的粘性末端可以连接成环 形分子。
pBR322质粒
pBR322质粒是由三个不同来源的部分组成的:第 一部分来源于pSF2124质粒的氨苄青霉素抗性基 因(ampr);
第二部分来源于pSC101质粒的四环素抗性基因 (tetr);
第三部分则来源于ColE1的派生质粒pMB1的DNA
复制起点(ori)。
27
PstI
ScaI
Ampr
HindIII BamHI
④ lacZ的a肽互补 1)a-肽( lacZ’ ):
b-半乳糖苷酶N端的一段氨基酸片断 (11-41氨基酸),该段基因序列连接到 pUC载体上。
受体菌基因组的b-半乳糖苷酶基因的 缺失a肽(氨基端有缺失),不能形成 活性酶,不能分解Xgal
37
⑤ 载体lacZ’与a互补
pUC质粒载体上的lacZ’ 编码a肽与这个 缺失突变的b-半乳糖苷酶“互补”,又 能分解Xgal。产生蓝色物质。
15
16
2. DNA 连接酶 3.1 DNA连接酶(ligase)的发现
从细菌DNA环化现象推测,必定存在一种能 把两条DNA双链连接到一起的酶。
DNA复制一定有断口。 17
3.2 DNA ligase的特点
1. 两种DNA连接酶
(1)大肠杆菌连接酶 只能连接粘性末端。
(2)T4噬菌体的连接酶 不但能连接粘性末端, 还能连接齐平末端。
基因克隆的酶学基础

医学课件ppt
9
医学课件ppt
数字表示在 不同寄主中 生长的噬菌 体的成斑率, 表示限制程 度。
10
二.限制和修饰作用的分子机制
1.大肠杆菌宿主细胞 K株,B 株 ,有各自的限制和修 饰系统。
1) 它们均有三个连续的基因位点控制,hsdR; hsdM; hsdS.
2) hsdR编码限制性核酸内切酶---识别DNA分子特定 位点,将双链DNA切断。(DNA分子转化细胞:受 体细胞去掉hsdR基因位点)
医学课件ppt
21
② 对称性 ③ 限制酶切后产生两个末端,末端结
构是5’-P和3’-OH
EcoRI 5’-G A A T T C-3’ 3’-C T T A A G-5’
医学课件ppt
22
(3)末端种类
① 3’-端突起,个数为2或4个核苷酸
Pst I 5’-CTGCAG-3’ 5’-CTGCA
医学课件ppt
15
(3)Ⅲ型酶
这类酶可识别特定碱基顺序,并在这一 顺序的3’端24~26bp处切开DNA,所以它 的切割位点也是没有特异性的。
医学课件ppt
16
二、限制性内切酶的特点 1、定义、命名
(1)定义
广义指上述三个系统中的限制酶; 狭义指II型限制酶。
(2)命名
限制酶由三部分构成,即菌种名、菌系编号、分离顺序。 例如:HindⅢ 前三个字母来自于菌种名称H. influenzae, “d”表示菌系为d型血清型(菌株号);“Ⅲ”表示分离 到的第三个限制酶。
EcoRI—Escherichia coli RI
医学课件ppt
17
2、限制酶的特点
(1)基本特点 在DNA双链的特异性识别序列部位,切割
基因克隆的四大要素(Four Elements for Gene Cloning)

将外源基因通过体外重组后导入受体细胞,使该基因能在受体细胞内复制、转录、翻译和表达,整个操作称为基因重组技术。
要实施该技术必须具备四大要素:工具酶、载体、基因和受体(宿主)细胞。
一、工具酶:基因工程的基本技术是人工进行基因的剪切、拼接、组合。
基因是一段具有一定功能的将外源基因通过体外重组后导入受体细胞,使该基因能在受体细胞内复制、转录、翻译和表达,整个操作称为基因重组技术。
要实施该技术必须具备四大要素:工具酶、载体、基因和受体(宿主)细胞。
一、工具酶:基因工程的基本技术是人工进行基因的剪切、拼接、组合。
基因是一段具有一定功能的DNA 分子,要把不同基因的DNA 线形分子片段准确地切出来,需要各种限制性核酸内切酶(restriction endonuclease);要把不同片段连接起来,需要DNA 连接酶(DNA ligase);要合成基因或其中的一个片段,需要DNA 聚合酶(DNA polymerase)等。
因此,酶是DNA 重组技术中必不可少的工具,基因工程中所用的酶统称为工具酶。
工具酶就其用途而言可分为三大类:限制性内切酶、连接酶和修饰酶,其中限制性内切酶为一大类酶(达上千种)。
基因重组正是利用了这些工具酶对DNA 分子进行一系列的酶催化反应,才得以在体外实现DNA 分子的切割和连接。
因此,工具酶的发现为基因操作提供了十分重要的技术基础。
首先重点介绍限制性内切酶(restriction endonucleases=restriction enzyme),其他酶在相关内容中再一一介绍。
从分子生物学发展历史看,核酸限制性内切酶的发现和应用对该学科发展所起的作用是难以估量的。
首先使外源基因在大肠杆菌中克隆的实验是在1973 年完成的,Stanley Cohen,Herbert Boyer(见补充资料2.1)正是利用了限制性内切酶这一分子手术刀才得以实现。
核酸限制性内切酶是原核生物中的一类能识别双链DNA 中特定碱基顺序的核酸水解酶。
生物化工学科硕士学位研究生培养方案

生物化工学科硕士学位研究生培养方案生物化工学科硕士学位研究生培养方案(081703)一、培养目标培养为社会主义现代化建设服务,德、智、体全面发展的生物化工高层次专门人才。
具体要求:1、较好地掌握马克思列宁主义、毛泽东思想的基本原理和邓小平理论、“三个代表”重要思想;树立辩证唯物主义和历史唯物主义世界观。
2、拥护党的基本路线,热爱祖国,遵纪守法,品行端正。
具有强烈的事业心和求实创新、团结协作的精神;献身科学事业,积极为社会主义现代化建设服务。
3、掌握本学科领域内坚实的基础理论和系统的专门知识,具有从事科学研究工作或独立承担技术性工作的能力;具有较宽的知识面和较强的适应性。
4、能较熟练地掌握一门外语。
5、具有健康的体魄和良好的心理素质。
二、研究方向1、生物化学加工研究生物质资源通过生物化学加工方法制取清洁能源、酶制剂、蛋白饲料、功能性食品等的基础理论和应用开发。
2、酶工程研究微生物酶的菌种筛选、微生物酶的合成、酶学性质、酶的分离提纯、酶的应用等。
3、生化分离技术运用生物反应与生物分离的原理开发和设计生物分子分离过程、优化生物分离过程。
4、微生物基因工程重组微生物的构建和基因表达技术;木质纤维降解酶类及其相关基因的分子生物学研究。
三、学习年限和时间安排全日制硕士研究生的学习年限一般为3年。
按课程学习与论文工作并重原则,课程学习累计1-1.5学年,论文工作量不少于1学年。
根据实际情况,经本人申请、导师同意、学校批准,可适当提前或延长一年,在职硕士可延长二年。
四、课程设置、学分要求和课程说明1、课程设置和学分要求课程设置分为学位课和非学位课,总学分要求不少于32学分(其中实践性环节2学分),其中学位课程18学分,非学位课程12学分。
对于同等学力和跨学位考的硕士生需补本科生课程,其成绩可减半登记学分,不占应学32学分的总学分。
讲课20学时为1学分,实验课30学时为1学分。
选修课程由指导教师和研究生根据专业培养方向要求,以及研究生原有基础、特长及专业爱好共同商定。
第三章 基因克隆的酶学基础-2

分子克隆工具酶及其应用
限制性内切酶—主要用于DNA分子的特异切割 DNA甲基化酶—用于DNA分子的甲基化 核酸酶—用于DNA和RNA的非特异性切割 核酸聚合酶—用于DNA和RNA的合成 核酸连接酶—用于DNA和RNA的连接 核酸末端修饰酶—用于DNA和RNA的末端修饰 其它酶类--用于生物细胞的破壁、转化、核酸纯 化、检测等。
甲基化酶与识别序列
受甲基化影响的限制酶b AluⅠ, BamHⅡ, Bsp1286 DdeⅠ,HgiAⅠ, NheⅠ, PstⅠ BamHⅠ, MspⅠ ClaⅠ, MboⅠ, TaqⅠ
EcoRⅠ MstⅠ, PstⅠ, PvuⅡ BanⅡ,BglⅠ,Bsp1286,BstXⅠ,HaeⅢ, MspⅠ,NaeⅠ,NcoⅠ,SacⅡ, Sau96Ⅰ AhaⅡ, FnuDⅡ, HhaⅠ AhaⅡ,AvaⅠ,AvaⅡ,HpaⅡ,ScrFⅠ HinfⅠ, HphⅠ, Sau3AⅠ BamHⅠ, MspⅠ AluⅠ, PstⅠ AluI, AvaⅠ, EcoRV, HinCⅡ, HinfⅠ, MboⅠ, TaqⅠ, XmnⅠ
三. 限制性内切酶的定义、命名
1. 定义:广义指上述三个系统中的限制酶;
狭义指II型限制酶。
2. 命名:限制酶由三部分构成,即菌种名、
菌系编号、分离顺序。 例如:HindⅢ 前三个字母来自于菌种 名称H. influenzae,“d”表示菌系为d型 血清型;“Ⅲ”表示分离到的第三个限制酶。
EcoRI—Escherichia coli RI HindⅢ—Haemophilus influensae d Ⅲ SacI (II)—Streptomyces achromagenes I (Ⅱ)
3、基因克隆的酶学基础-01

7、完全酶切和部分酶切
3.2 DNA连接酶与DNA 分子的体外连接 3.2.1 DNA连接酶:大肠杆菌DNA连接酶和T4DNA连接酶
动物细胞和噬菌体
缺口(nick)
裂口(gap)
大肠杆菌及其它细菌
3.2.2 粘性末端的连接
用碱性磷酸 酶的脱磷酸 作用阻止线
S1 核酸酶在分子生物学研究中的主要用途是给RNA定位。
3.6.2 Bal 31 核酸酶与限制位点的确定 具有单链特异的核酸内切酶活性和双链特异的核酸外切酶活性。
应用Bal 31 核酸酶诱发 DNA分子的缺失突变
(a)质粒DNA分子,A区段是准 备诱发缺失突变的序列
(b)用Bal 31核酸酶不同时间长 度分别消化线性DNA分子 S1 核酸酶与RNA分子定位 反应条件:低水平Zn2+、最适pH值:4.0~4.3、NaCI浓度:
100mmol/L
S1 核酸酶的主要功能: ① 催化RNA和单链DNA分子降解成5′单核苷酸; ② 作用于双链核酸分子的单链区,并从此处切断核酸分子 (单链区可小到一个碱基对)。
T4 多核苷酸激酶的交换活性
3.4.2 碱性磷酸酶(BAP、CIP)与 DNA脱磷酸作用
3.5 DNA 外切酶
核酸外切酶(exonucleases):是一类从多核苷酸链的一 头开始按序催化降解核苷酸的酶。按作用特性的差异可分为 单链核酸外切酶(如exo I、 exo VII等 )和双链核酸外切酶 (如exo III、λ exo等)。
(c)将带有另一种限制性内切酶 识别序列的衔接物连接到这些缺 失分子上并将它们环化起来
(d)新形成的环状质粒分子具有 新的限制性内切酶(Hind III)的 切点,其A区段有缺失
8-1-基因克隆的工具酶

9
2.1 DNA聚合酶Ⅰ (DNApolymerase I)
5ˊ→3ˊ聚合酶活性 3ˊ→5ˊ核酸外切酶活性 5ˊ→3ˊ核酸外切酶活性
10
2.2 DNA聚合酶Ⅰ大片段
(large fragment of DNApolymerase I)
DNA聚合酶I用枯草杆菌蛋白酶(subtilisin)裂解后产生的大 片段,这个片段也称为Klenow片段(Klenow fragment)。 5ˊ→3ˊ聚合酶活性 3ˊ→5ˊ核酸外切酶活性 无5ˊ→3ˊ核酸外切酶活性
限制性内切酶的命名
EcoR Ⅰ: E代表Escherichia属 co代表coli 种 R代表RY13株
7
限制性内切酶的识别和切割位点
通常是4~6个碱基对、具有回文序列(palindrom)的 DNA片段,大多数酶错位切割双链DNA,产生5ˊ或3ˊ黏性末 端(sticky end)。
8
2. 其他常用工具酶
第八章 基因克隆与体外表达
基因克隆的工具酶
1
。 Paul Berg:1926~ Stanley N. Cohen
自从1973年首次采用质粒与外源 DNA体外重组的方法克隆 DNA以来,重组DNA技术作为分子生物学的一项重要技术得到了迅速 发展,使科学家们分离、分析及操作基因的能力达到几乎无所不能的地步。
活性 识别特异碱基序列,切割DNA 催化DNA5ˊ-磷酸与3ˊ-羟基形成磷酸二酯键
DNA聚合酶 逆转录酶 碱性磷酸酶 末端脱氧核苷酸转移酶
以DNA为模板合成DNA 以RNA为模板合成cDNA 切除5端磷酸根 催化3端合成同聚尾
18
思考题:
简述限制性核酸内切酶等基因克隆 常用工具酶的异同点,并说说它们在基 因克隆操作中的重要性。
基因工程习题题目练习(附答案)版

基因⼯程习题题⽬练习(附答案)版基因⼯程原理复习题思考题考试时间:2009.06.21 上午9:00-11:00 地点5D305基因⼯程绪论1、基因⼯程的定义与特征。
定义:在体外把核酸分⼦(DNA的分离、合成)插⼊载体分⼦,构成遗传物质的新组合(重组DNA),引⼊原先没有这类分⼦的受体细胞内,稳定地复制表达繁殖,培育符合⼈们需要的新品种(品系),⽣产⼈类急需的药品、⾷品、⼯业品等。
特征:1、具跨越天然物种屏障的能⼒。
2、强调了确定的DNA⽚段在新寄主细胞中的扩增。
2、试述基因⼯程的主要研究内容。
1)、⽬的基因的分离2)、DNA的体外重组(载体、受体系统等)3)、重组DNA分⼦转移到受体细胞及其筛选4)、基因在受体细胞内的扩增、表达、检测及其分析。
3、基因⼯程在⾷品⼯业上有何应⽤发展?主要是通过基因重组,使各种转基因⽣物提⾼⽣产⾕氨酸、调味剂、酒类和油类等有机物的产率;或者改良这些有机物组成成分,提⾼利⽤价值。
4、转基因是⼀把双刃剑,请客观谈谈对转基因及转基因⾷品安全性的认识。
转基因技术所带来的好处是显⽽易见的,在⼈类历史进步和发展中起到了积极作⽤。
⾸先,通过该项技术可以提供⼈们所需要的特性,改良培育新品种;第⼆,延长⾷品保存时间或增加营养成分;第三,将抗⾍防菌基因转⼊到作物中,使作物本⾝产⽣抵抗病⾍害侵袭的能⼒,减少了农药的使⽤量,有利于环境保护;第四,转基因技术及基因⾷物在医学⽅⾯得到⼴泛研究和应⽤。
⼈们对转基因技术的主要担忧在于环境⽅⾯。
外源基因的导⼊可能会造就某种强势⽣物,产⽣新物种或超级杂草、损害⾮⽬标⽣物、破坏原有⽣物种群的动态平衡和⽣物多样性,也即转基因⽣物存在潜在的环境安全问题。
转基因作物的⼤⾯积种植已有数年,⾷⽤转基因⾷品的⼈群⾄少有10亿之多,但⾄今仍未有转基因⾷品对⽣命造成危害的实例;更何况⽬前每⼀种基因⼯程⾷品在上市前,都要经过国家法律认可,⾷品卫⽣部门和环境部门的严格检测。
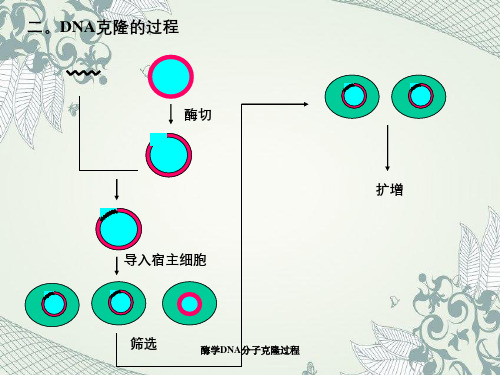
酶学DNA分子克隆过程
NheI GCTAGC CGATCG
XhoI CTCGAG GAGCTC
先用NheI切割
CTAGC G
用T4DNA聚合酶或 Klenow酶补平
CTCGAG GAGCTC dNTP
CTAGC GATCG XhoI切割
CTCGAG GAGCTC
酶学DNA分子克隆过程
CTAGC GATCG
C GAGCT
之间的连接。
7。DNA连接酶
活性:催化双链DNA中相邻碱基的5‘磷酸和3’羟基间磷酸二酯 键的形成。
(1)T4 DNA连接酶:
此酶是唯一在正常条件下,有效连接平端DNA的连接酶。粘性 末端的连接通常在12~15度进行,平端的连接通常在室温 (低于30度),所需酶酶学D量NA是分子粘克端隆连过程接的10~100倍。
3,两端都不匹配
(1)平末端连接 NheI
GCTAGC CGATCG
用NdeI和NheI切割,回
NdeI CATATG GTATAC
酶学DNA分子克隆过程
G
载体
CTAGC G
用T4DNA聚合酶补平 CTAGC GATCG
CA GTAT
dNTP
CATA GTAT
用碱性磷酸酶去除 5端磷酸根
酶学DNA分子克隆过程
用途:
(1)
用于DNA测序
(2) 用于标记5‘末端突出的DNA的3’末端。由于它会在3‘端
留下一个额外的碱基,因此不能应用于将粘性DNA末端
转变成平端。
4。 反转录酶
来源:
1. 禽类成髓细胞瘤病毒 (avian myoblastosis virus,AMV) 2. 莫洛尼鼠白血病病毒 (Moloney murine leukemia virus,MMLV)
基因克隆的酶学基础

基因工程
第三章 基因工程酶学基础
返回第三章
2,产生克隆位点 两个不同的DNA样品用同一种酶酶切后,能产生相同的粘性末端
两个平末端DNA片段也可以进行相互拼接
HindIII --CTCGAAGCTTGTTC--CAGCTTCGAACAAG--CTCA—p HO—ACAAG混合 退火
DNA编码,对平末端的连接效率比细菌DNA连接酶高的多。
热稳定的DNA连接酶:嗜热高温放线菌中分离,85度有高活性,
94仍有活性。
西北师范大学 精品课程 武国凡
基因工程
第三章 基因工程酶学基础
返回第三章
3.2.3 连接酶的连接作用
西北师范大学 精品课程 武国凡
基因工程
第三章 基因工程酶学基础
返回第三章
西北师范大学 精品课程 武国凡
基因工程
第三章 基因工程酶学基础
西北师范大学 精品课程 武国凡
基因工程
第三章 基因工程酶学基础
返回第三章
3.1.6 Ⅱ型酶的特点和识别位点
•大多数II型酶结合并且切割回文序列,又称为识别序列或识别位点
•酶切后产生5’磷酸突出或3’羟基突出的末端,它们统称为粘性末端 酶 •有一些在切割两条链时会产生两端平整(平末端)的DNA分子 •酶的识别位点的可以是4个、5个、6个、8个甚至更多的碱基对 •在分子克隆实验中使用最普遍的是识别4个或6个碱基对的酶 •星号活性:非最佳条件下表现出的活性
西北师范大学 精品课程 武国凡
基因工程
第三章 基因工程酶学基础
返回第三章
第二节 DNA、RNA连接酶 3.2.1 待 连 接 二 端 之 一 须 有 磷 酸 根 连接酶
西北师范大学 精品课程 武国凡
- 1、下载文档前请自行甄别文档内容的完整性,平台不提供额外的编辑、内容补充、找答案等附加服务。
- 2、"仅部分预览"的文档,不可在线预览部分如存在完整性等问题,可反馈申请退款(可完整预览的文档不适用该条件!)。
- 3、如文档侵犯您的权益,请联系客服反馈,我们会尽快为您处理(人工客服工作时间:9:00-18:30)。
b .3’一侧切割形成3’突出末端
5’-CTGCAG-3’ 3’-GACGTC-5’
PstI切割位点
PstI等产生的3‘粘性末端
5‘ G-C-T-C-T-G-C-A-G-G-A-G 3’ 3‘ C-G-A-G-A-C-G-T-C-C-T-C 5’
PstI 37
5‘ … G-C-℃T-C-T-G-C-A-OH P-G-G-A-G … 3’ 3‘ … C-G-A-G-P OH-A-C-G-T-C-C-T-C … 5’
第四章 分子克隆的酶学基础
基因的重组与分离,涉及到一系列相互关 联的酶促反应。特别是核酸限制性内切酶 和 DNA连接酶的发现和应用,使DNA分子的体外 连接与切割成为可能。
核酸限制性内切酶和DNA连接酶是重组 DNA技术赖以创立的重要酶学基础。
重组DNA实验中常使用的酶
① II型核酸内切限制酶 ② DNA连接酶 ③ 大肠杆菌DNA 聚合酶 ④ 反转录酶 ⑤ 多核苷酸激酶 ⑥ 末端转移酶
用其它二价阳离子如 Mn++ 、Cu++、Co++ 或 Zn++ 代替了 Mg++ 。
10、 酶切位点的引入
(1)产生的 5‘ 突出端补平后,再连接可产 生新的酶切位点
(2)同尾末端的连接
不同的同尾酶切割 DNA 产生的末端再相互连接时,可产生新的 酶切位点,同时原来的酶切位点消失
BamHⅠ(G↓GATCC)+ BclⅠ(T↓GATCA)
EcoB
EcoK
修饰的甲基转移酶 核酸内切限制酶
EcoR I
GAATTC CTTAAG
M EcoR I
限制作用
G3’ 5’ AATTC CTTAA5’ 3’ G
EcoR I
修饰作用
Me GAATTC CTTAAG
Me
核酸内切限制酶EcoR I 及其修饰的甲基化酶MEcoR 的限制与修饰作用
寄主的限制与修饰的作用
增加核酸内切酶的用量,1ugDNA/10u; 扩大反应体积,稀释抑制因素 延长酶切保温时间
(2)甲基化程度:大肠杆菌中的dam甲基化酶在 5‘GATC3’序列中的腺嘌呤N6位引入甲基
(3)缓冲液
pH,Mg2+ ,DTT,BSA,10(X)
(4) 酶切消化反应的温度
大多数为 37℃,少数 25-30℃
P-C-T-G-G-A-G … 3’ OH-G-A-C-C-T-C … 5’
同裂酶(Isoschizomer): 识别位点的序列相同的限制性 内切酶
完全同裂酶:识别位点和切点完全相同:如MobI和Sau3AI 识别序 列切割位点为 GATC
不完全同裂酶:识别位点相同,但切点不同如AatII(GACGT ↓ C )ZraI(GAC ↓ GTC)
因此
设计 PCR 引物时,引入酶切位点时,在位点之外加上 能够满足要求的碱基数目。
双酶切多克隆位点时选择合适的酶切秩序
7、位点偏爱(site preference)
某些限制酶对同一介质中的有些位点表现出偏爱性切 割,即对不同位置的同一个识别序列表现出不同的 切割效率的现象称作位点偏爱
(1) 现象:
噬菌体 DNA 有 4 个 SacⅡ 位点,三个在中央,一个在右臂,对 中央三个位点的酶切速度快 50 倍
8.II 型核酸内切酶的多酶联合酶解:
对盐浓度要求相同的酶,原则上可以同时酶切, 对盐浓度要求不同的酶,可采取下列方法: 使用较贵的酶的盐浓度,加大便宜酶的用量,同时
酶解 低盐酶先切,然后补加盐,高盐酶再切
EcoRⅠ 酶切割 噬菌体中的 5 个位点时并不是随机的,靠近右端 的位点比分子中间的位点切割快 10 倍;
EcoRⅠ 对腺病毒 2(adenvirus-2)DNA 不同位置切点的切割速 率也不同。
EcoRⅠ 和 HindⅢ 在 噬菌体 DNA 中的切割速率分别有 10 倍 和 14 倍的差异
2006年2月为止共发现3773种限制酶,810种 甲基化酶
核酸内切限制酶 (restriction endonuclease)
是一类能够识别双链DNA分子中的某种特定 核苷酸序列(4-8bp),并由此处切割DNA 双链的核酸内切酶
3. 核酸内切限制酶的类型
特性
蛋白质结构 辅助因子 序列特异切割
① 在特异识别序列切割DNA分子; ② 两个单链切割部位在DNA分子上的分布,通常不直接相对; ③ 断片往往具有互补的单链延伸末端。
识别序列
4-8个核苷酸序列
①大部分呈旋转对称、反向重复结构—回文结构
切割位点
C-C-G-C-G-G G-G-C-G-C-C
切割位点 对称轴
A B C C΄ B΄ A΄ A΄B΄ C΄ C B A
甘油浓度高(>5%)
酶过量(>100U/l)
离子强度低(<25mM)
pH 值过高(>8.0)
有机溶剂如 PMSD (二甲基亚砜)、乙醇、乙二醇、二甲基 乙酰胺(dimethylacetamide)、二甲基甲酰胺 (dimethbylformamide)、 sulphalane 等等.
5‘
…
OH
G-C-T-C-T-G-C-A
P
G-G-A-G
…
3’
3‘ … C-G-A-G A-C-G-T-C-C-T-C … 5’
P OH
5’突出端易于通过DNA激酶和32p ATP进 行同位素标记
3’突出端是末端转移酶的理想作用底物,在 酶的作用下,容易使DNA片段带上多核苷酸 尾
平末端 Blunt ends
A B B΄ A΄ A΄ B΄ A B
② 部分识别序列不对称:
AccBSⅠ :CCG↓CTC GGC↑GAG
BssSⅠ : C↓TCGTG GAGCA↑C
③ 有一些限制酶可识别多种序列 AccI GT MKAC (M:A或C)
HindII GTY RAC(Y:C或R:A或G)
识别位点在DNA分子中的频率
从核酸分子末端开始,一个核苷酸一个核 苷酸地消化降解多核苷酸链,称为核酸外 切酶( exonuclease);从核酸内部切割 磷酸二酯键使之断裂为小片段的为核酸内 切酶(endonuclease)。
第一节、核酸内切限制酶与DNA分子的体外切割
1. 寄主控制的限制与修饰现象(R/M)
(Restriction and modification)
a .在对称轴的5’一侧切割底物, DNA双链交错断开,形成5‘突出末端
5’-GAATTC-3’ 3’-CTTAAG-5’
EcoRI切割位点
EcoRI等产生的5‘粘性末端
5‘ … G-C-T-G-A-A-T-T-C-G-A-G … 3’ 3‘ … C-G-A-C-T-T-A-A-G-C-T-C … 5’
切割位点
I型酶
II型酶
3种亚基
单一成分
ATP, Mg2+, S-Met
Mg2+
不是Байду номын сангаас
是
距识别序列1kb 识别序列内 处
随机切割 或附近
III型酶
2种不同亚基 ATP, Mg2+, S-Met
是
距识别距离下游 24-26bp处
克隆中的作用
无用
十分有用
用处不大
4. II型核酸内切限制酶的基本特性
(1) 基本特性
EcoRI 37 ℃
5‘ … G-C-T-G-OH
P-A-A-T-T-C-G-A-G … 3’
3‘ … C-G-A-C-T-T-A-A-P
OH-G-C-T-C …
5’
退火 4-7 ℃
OH P
5‘ … G-C-T-G A-A-T-T-C-G-A-G … 3’
3‘ … C-G-A-C-T-T-A-A G-C-T-C … 5’
(2)、 限制性内切酶产生的末端
粘性末端Cohesive ends (Mached ends)
5‘粘性末端, 3’突出末端
平末端 Blunt ends 非对称突出端
①粘性末端Cohesive ends (Mached ends)
DNA分子在限制性内切酶的作用下形成具有互补碱基的单 链延伸末端结构,能够通过互补碱基的配对而重新连接起来.
假定核苷酸随机排列的情况下进行
44=256 46=4096
实践中: 如λ DNA长49kb,对于六碱基的酶应该有12个切割位点,
实际上要少一些,如Bgl II只有6个,BamHI只有5个, 而SalI只有2个。
基因组中碱基对的排列是非均匀的 ,尽管有的酶 切序列中 GC 含量相同,但在基因组中出现的
具粘性末端的DNA片段结合方式
6. 核酸内切限制酶的命名法
① 属名头一个字母+种名头两个字母 ② 菌株名 ③ 不同修饰体系 ④ 系统名
Haemophilus influenzae d 嗜血流感杆菌d株 HindIII
DNA 片段(线性载体)检测末端长度对切割的影响时,同 样发现识别序列的末端长度对酶切效率有明显影响,不 同的酶对末端长度的要求是不同。
性
星星活性是限制性内切酶的一般性质,任何一种限 制酶在极端非标准条件下都能切割非典型位点
ApoⅠ、AseⅠ、BamHⅠ、BssH11、EcoRⅠ、 EcoRV、HindⅢ、HinfⅠ、KpnⅠ、PstⅠ、 PvuⅡ、SalⅠ、ScaⅠ、TaqⅠ 和 XmnⅠ 等酶 皆可表现出星星活性。
高浓度的酶、高浓度的甘油、低离子强度、极端pH等会使一核酸 内切酶的识别和切割序列发生低特异性,即所谓的Star activity现象
smalI(25℃);ApaI(30℃)
(5)DNA的分子构型